

Freud-1: A neuronal calcium-regulated repressor of the 5-HT1A receptor gene
- PMID: 12917378
- PMCID: PMC6740452
- DOI: 10.1523/JNEUROSCI.23-19-07415.2003
Freud-1: A neuronal calcium-regulated repressor of the 5-HT1A receptor gene
Abstract
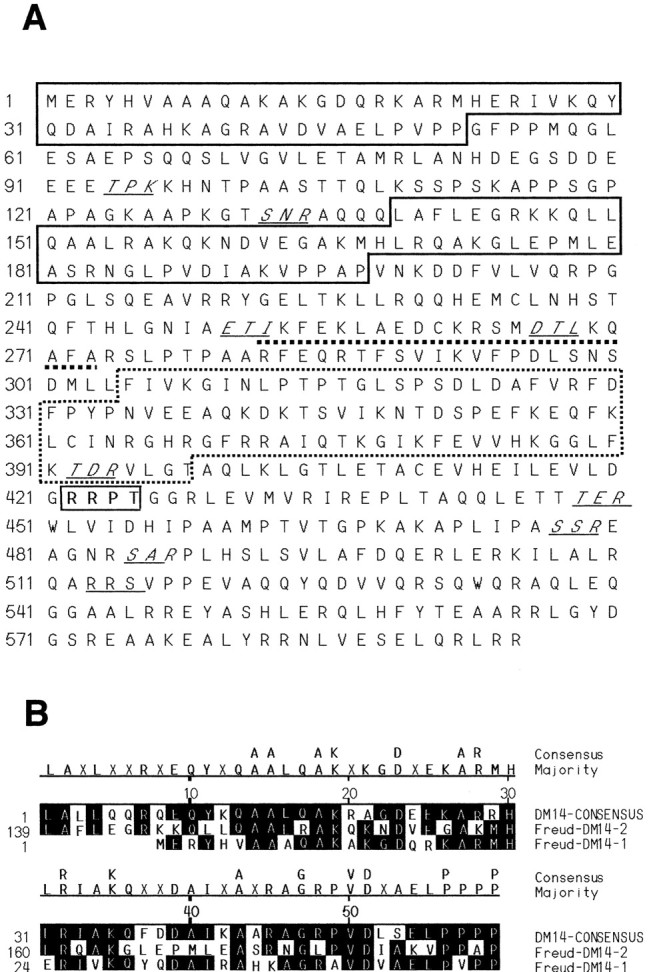
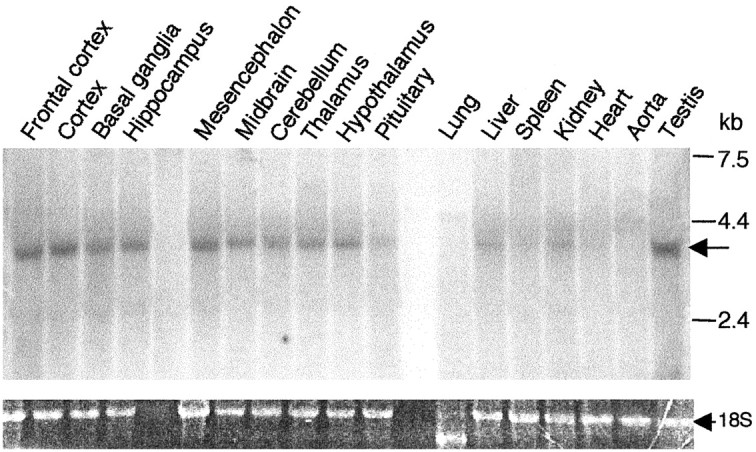
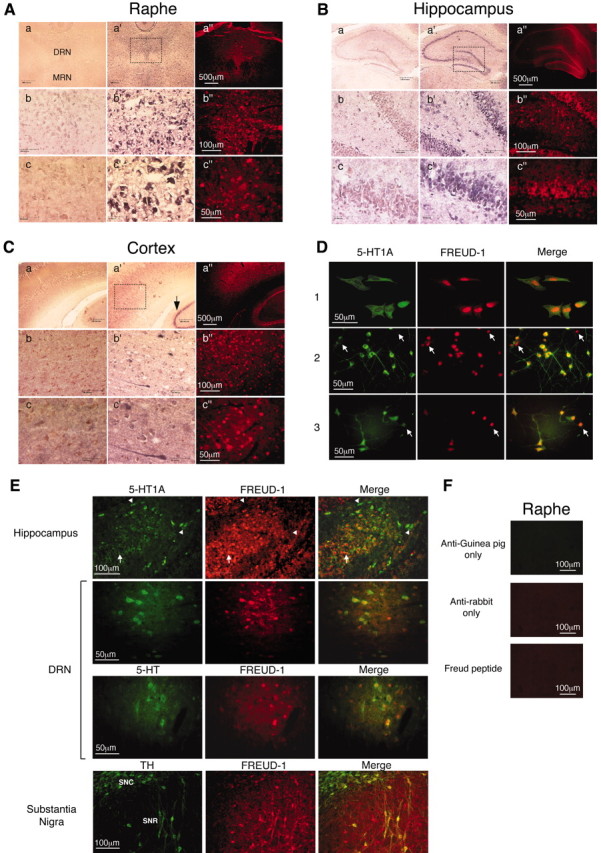
Altered regulation of 5-HT1A receptors is implicated in mood disorders such as anxiety and major depression. To provide insight into its transcriptional regulation, we previously identified a novel DNA element [14 bp 5'-repressor element (FRE)] of the 5-HT1A receptor gene that mediates repression in neuronal and non-neuronal cells (Ou et al., 2000). We have now cloned a novel DNA binding protein [five' repressor element under dual repression binding protein-1 (Freud-1)] that binds to FRE to mediate repression of the 5-HT1A receptor or heterologous promoters. Freud-1 is evolutionarily conserved and contains two DM-14 basic repeats, a predicted helix-loop-helix DNA binding domain, and a protein kinase C conserved region 2 (C2)/calcium-dependent lipid binding (CalB) calcium/phospholipid binding domain. An intact CalB domain was required for Freud-1-mediated repression. In serotonergic raphe cells, overexpression of Freud-1 repressed the 5-HT1A promoter and decreased 5-HT1A receptor protein levels, whereas transfection of antisense to Freud-1 derepressed the 5-HT1A gene and increased 5-HT1A receptor protein expression. Calcium-dependent signaling blocked Freud-1-FRE binding and derepressed the 5-HT1A promoter. Treatment with inhibitors of calmodulin or CAM-dependent protein kinase reversed calcium-mediated inhibition of Freud-1. Freud-1 RNA and protein were present in raphe nuclei, hippocampus, cortex, and hypothalamus, and Freud-1 protein was colocalized with 5-HT1A receptors, suggesting its importance in regulating 5-HT1A receptors in vivo. Thus, Freud-1 represents a novel calcium-regulated repressor that negatively regulates basal 5-HT1A receptor expression in neurons and may play a role in the altered regulation of 5-HT1A receptors associated with anxiety or major depression.
Figures



References
-
- Abdouh M, Storring JM, Riad M, Paquette Y, Albert PR, Drobetsky E, Kouassi E ( 2001) Transcriptional mechanisms for induction of 5-HT1A receptor mRNA and protein in activated B and T lymphocytes. J Biol Chem 276: 4382-4388. - PubMed
-
- Albert PR, Tashjian Jr AH ( 1986) Ionomycin acts as an ionophore to release TRH-regulated Ca 2+ stores from GH4C1 cells. Am J Physiol 251: C887-891. - PubMed
-
- Albert PR, Zhou QY, Van Tol HH, Bunzow JR, Civelli O ( 1990) Cloning, functional expression, and mRNA tissue distribution of the rat 5-hydroxy-tryptamine1A receptor gene. J Biol Chem 265: 5825-5832. - PubMed
-
- Albert PR, Lembo P, Storring JM, Charest A, Saucier C ( 1996) The 5-HT1A receptor: signaling, desensitization, and gene transcription. Neuropsychopharmacology 14: 19-25. - PubMed
-
- Albert PR, Sajedi N, Lemonde S, Ghahremani MH ( 1999) Constitutive G(i2)-dependent activation of adenylyl cyclase type II by the 5-HT1A receptor. Inhibition by anxiolytic partial agonists. J Biol Chem 274: 35469-35474. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous