

Regulation of expression of scaffoldin-related genes in Clostridium thermocellum
- PMID: 12923083
- PMCID: PMC181014
- DOI: 10.1128/JB.185.17.5109-5116.2003
Regulation of expression of scaffoldin-related genes in Clostridium thermocellum
Abstract
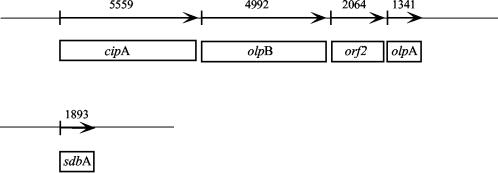
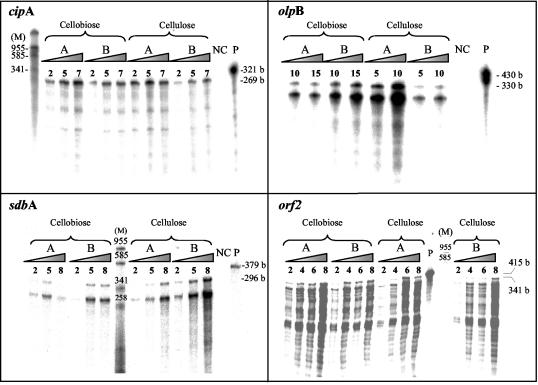
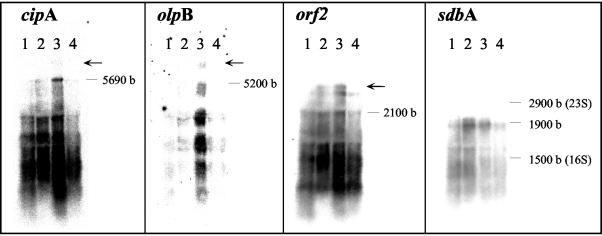
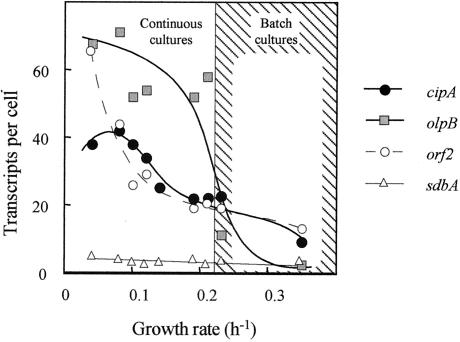
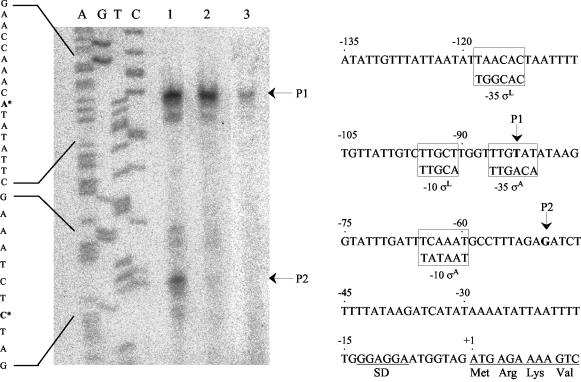
Clostridium thermocellum produces an extracellular multienzyme complex, termed the cellulosome, that allows efficient solubilization of crystalline cellulose. The complex is organized around a large noncatalytic protein subunit, termed CipA or scaffoldin, and is found either free in the supernatant or cell bound. The binding of the complex to the cell is mediated by three cell surface anchoring proteins, OlpB, Orf2p, and SdbA, that interact with the CipA scaffoldin. The transcriptional level of the olpB, orf2, sdbA, and cipA genes was determined quantitatively by RNase protection assays in batch and continuous cultures, under carbon and nitrogen limitation. The mRNA level of olpB, orf2, and cipA varied with growth rate, reaching 40 to 60 transcripts per cell under carbon limitation at a low growth rate of 0.04 h(-1) and 2 to 10 transcripts per cell at a growth rate of 0.35 h(-1) in batch culture. The mRNA level of sdbA was about three transcripts per cell and was not influenced by growth rate. Primer extension analysis revealed two major transcriptional start sites, at -81 and -50 bp, upstream of the translational start site of the cipA gene. The potential promoters exhibited homology to the known sigma factors sigma(A) and sigma(L) (sigma(54)) of Bacillus subtilis. Transcription from the sigma(L)-like promoter was found under all growth conditions, whereas transcription from the sigma(A)-like promoter was significant only under carbon limitation. The overall expression level obtained in the primer extension analysis was in good agreement with the results of the RNase-protection assays.
Figures
References
-
- Bayer, E. A., E. Morag, Y. Shoham, J. Tormo, and R. Lamed. 1996. The cellulosome: a cell-surface organelle for the adhesion to and degradation of cellulose, p. 155-182. In M. Fletcher (ed.), Bacterial adhesion: molecular and ecological diversity. Wiley-Liss, Inc., New York, N.Y.
-
- Bayer, E. A., L. J. Shimon, Y. Shoham, and R. Lamed. 1998. Cellulosomes: structure and ultrastructure. J. Struct. Biol. 124:221-234. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
