

Cell signal control of the G protein-gated potassium channel and its subcellular localization
- PMID: 12923211
- PMCID: PMC1664760
- DOI: 10.1113/jphysiol.2003.048439
Cell signal control of the G protein-gated potassium channel and its subcellular localization
Abstract
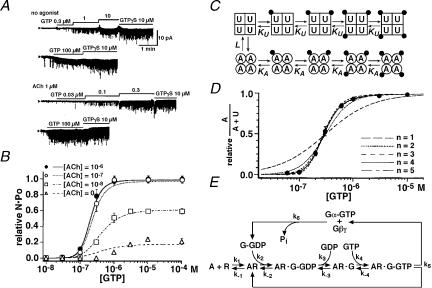
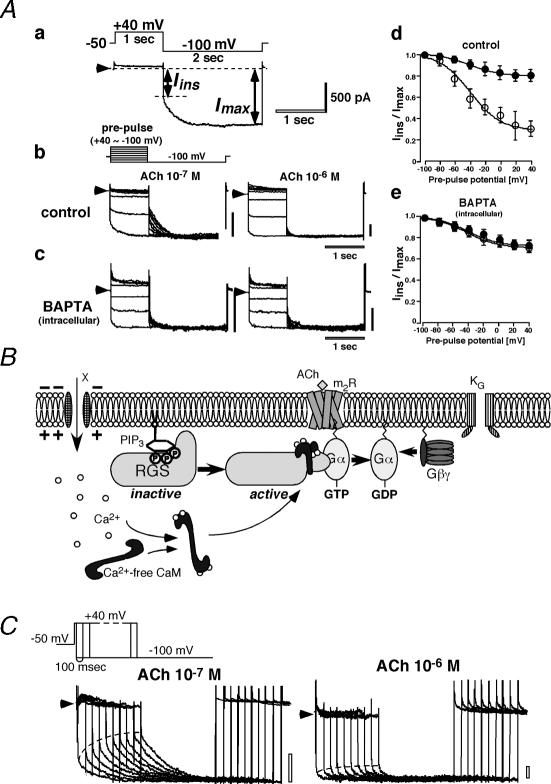
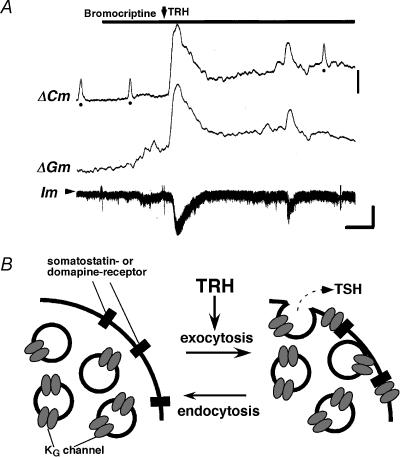
G protein-gated inward rectifier K(+) (K(G)) channels are directly activated by the betagamma subunits released from pertussis toxin-sensitive G proteins, and contribute to neurotransmitter-induced deceleration of heart beat, formation of slow inhibitory postsynaptic potentials in neurones and inhibition of hormone release in endocrine cells. The physiological roles of K(G) channels are critically determined by mechanisms which regulate their activity and their subcellular localization. K(G) channels are tetramers of inward rectifier K(+) (Kir) channel subunits, Kir3.x. The combination of Kir3.x subunits in each K(G) channel varies among tissues and cell types. Each subunit of the channel possesses one Gbetagamma binding site. The binding of Gbetagamma increases the number of functional K(G) channels via a mechanism that can be described by the Monod-Wyman-Changeux allosteric model. During voltage pulses K(G) channel current alters time dependently. The K(G) current exhibits inward rectification due to blockade of outward-going current by intracellular Mg(2+) and polyamines. Upon repolarization, this blockade is relieved practically instantaneously and then the current slowly increases further. This slow current alteration is called 'relaxation'. Relaxation is caused by the voltage-dependent behaviour of regulators of G protein signalling (RGS proteins), which accelerate intrinsic GTP hydrolysis mediated by the Galpha subunit. Thus, the relaxation behaviour of K(G) channels reflects the time course with which the G protein cycle is altered by RGS protein activity at each membrane potential. Subcellular localization of K(G) channels is controlled by several distinct mechanisms, some of which have been recently clarified. The neuronal K(G) channel, which contains Kir3.2c, is localized in the postsynaptic density (PSD) of various neurones including dopaminergic neurones in substantia nigra. Its localization at PSD may be controlled by PDZ domain-containing anchoring proteins. The K(G) channel in thyrotrophs is localized exclusively on secretary vesicles, which upon stimulation are rapidly inserted into the plasma membrane and causes hyperpolarization of the cell. This mechanism indicates a novel negative feedback regulation of exocytosis. In conclusion, K(G) channels are under the control of a variety of signalling molecules which regulate channel activity, subcellular localization and thus their physiological roles in myocytes, neurones and endocrine cells.
Figures

Similar articles
-
Interaction between the RGS domain of RGS4 with G protein alpha subunits mediates the voltage-dependent relaxation of the G protein-gated potassium channel.J Physiol. 2001 Aug 15;535(Pt 1):133-43. doi: 10.1111/j.1469-7793.2001.t01-1-00133.x. J Physiol. 2001. PMID: 11507164 Free PMC article.
-
Characterization of G-protein-gated K+ channels composed of Kir3.2 subunits in dopaminergic neurons of the substantia nigra.J Neurosci. 1999 Feb 1;19(3):1006-17. doi: 10.1523/JNEUROSCI.19-03-01006.1999. J Neurosci. 1999. PMID: 9920664 Free PMC article.
-
Agonist unbinding from receptor dictates the nature of deactivation kinetics of G protein-gated K+ channels.Proc Natl Acad Sci U S A. 2003 May 13;100(10):6239-44. doi: 10.1073/pnas.1037595100. Epub 2003 Apr 28. Proc Natl Acad Sci U S A. 2003. PMID: 12719528 Free PMC article.
-
Inwardly rectifying potassium channels: their structure, function, and physiological roles.Physiol Rev. 2010 Jan;90(1):291-366. doi: 10.1152/physrev.00021.2009. Physiol Rev. 2010. PMID: 20086079 Review.
-
Pharmacology of cardiac potassium channels.Adv Pharmacol. 2010;59:93-134. doi: 10.1016/S1054-3589(10)59004-5. Adv Pharmacol. 2010. PMID: 20933200 Review.
Cited by
-
Increased GIRK channel activity prevents arrhythmia in mice with heart failure by enhancing ventricular repolarization.Sci Rep. 2023 Dec 18;13(1):22479. doi: 10.1038/s41598-023-50088-2. Sci Rep. 2023. PMID: 38110503 Free PMC article.
-
Minireview: potassium channels and aldosterone dysregulation: is primary aldosteronism a potassium channelopathy?Endocrinology. 2014 Jan;155(1):47-55. doi: 10.1210/en.2013-1733. Epub 2013 Dec 20. Endocrinology. 2014. PMID: 24248457 Free PMC article. Review.
-
A Quantitative Model of the GIRK1/2 Channel Reveals That Its Basal and Evoked Activities Are Controlled by Unequal Stoichiometry of Gα and Gβγ.PLoS Comput Biol. 2015 Nov 6;11(11):e1004598. doi: 10.1371/journal.pcbi.1004598. eCollection 2015 Nov. PLoS Comput Biol. 2015. PMID: 26544551 Free PMC article.
-
Dopaminergic intracellular signal integrating proteins: relevance to schizophrenia.Dialogues Clin Neurosci. 2006;8(1):95-100. doi: 10.31887/DCNS.2006.8.1/bsouza. Dialogues Clin Neurosci. 2006. PMID: 16640119 Free PMC article. Review.
-
Molecular and cellular diversity of neuronal G-protein-gated potassium channels.J Neurosci. 2005 Dec 7;25(49):11468-78. doi: 10.1523/JNEUROSCI.3484-05.2005. J Neurosci. 2005. PMID: 16339040 Free PMC article.
References
-
- Breitwieser GE, Szabo G. Uncoupling of cardiac muscarinic and β-adrenergic receptors from ion channels by a guanine nucleotide analogue. Nature. 1985;317:538–540. - PubMed
-
- Cohen NA, Brenman JE, Snyder SH, Bredt DS. Binding of the inward rectifier K+ channel Kir 2.3 to PSD-95 is regulated by protein kinase A phosphorylation. Neuron. 1996;17:759–767. - PubMed
-
- Corey S, Clapham DE. The Stoichiometry of Gβγ binding to G-protein-regulated inwardly rectifying K+ channels (GIRKs) J Biol Chem. 2001;276:11409–11413. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources