

Estrogen deficiency induces bone loss by increasing T cell proliferation and lifespan through IFN-gamma-induced class II transactivator
- PMID: 12923292
- PMCID: PMC193574
- DOI: 10.1073/pnas.1533207100
Estrogen deficiency induces bone loss by increasing T cell proliferation and lifespan through IFN-gamma-induced class II transactivator
Abstract
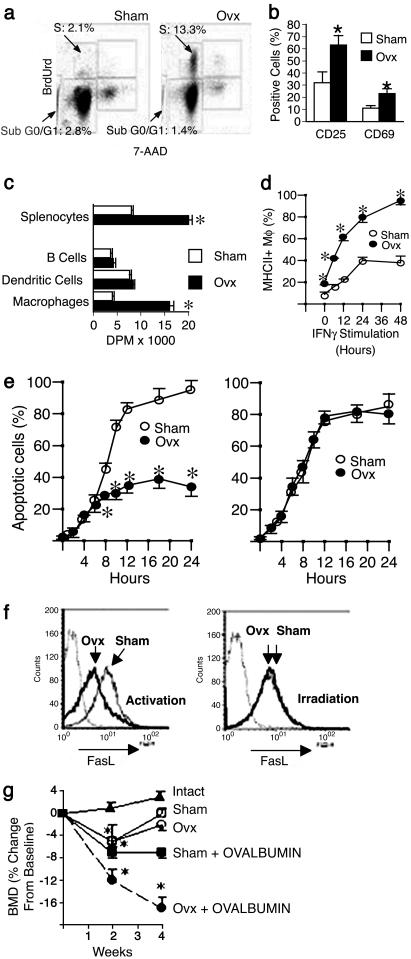
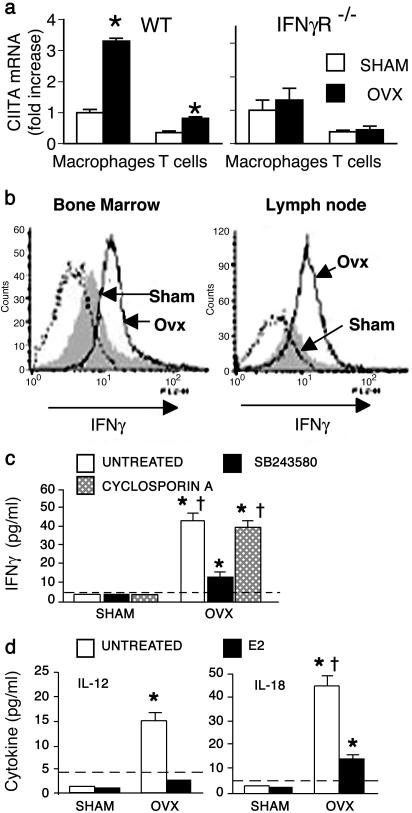
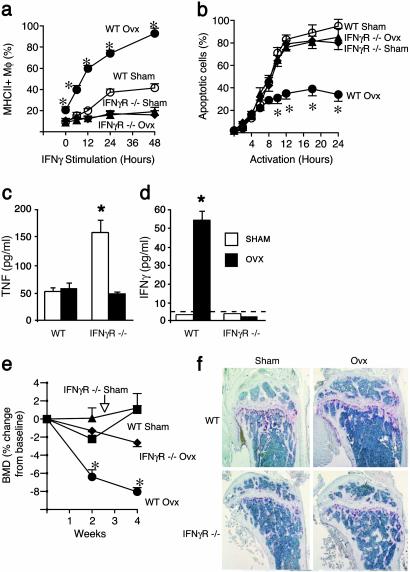
Expansion of the pool of tumor necrosis factor (TNF)-alpha-producing T cells is instrumental for the bone loss induced by estrogen deficiency, but the responsible mechanism is unknown. Here we show that ovariectomy up-regulates IFN-gamma-induced class II transactivator, a multitarget immune modulator, resulting in increased antigen presentation by macrophages, enhanced T cell activation, and prolonged lifespan of active T cells. Up-regulation of class II transactivator derives from increased production of IFN-gamma by T helper 1 cells, resulting from enhanced secretion of IL-12 and IL-18 by macrophages. The resulting T cell expansion and bone loss are prevented in vivo by both blockade of antigen presenting cell-induced T cell activation, and silencing of IFN-gamma receptor signaling. Thus, increased IFN-gamma-induced class II transactivator expression and the resulting enhanced T cell proliferation and lifespan are critical to the bone wasting effect of estrogen deficiency.
Figures
References
-
- Riggs, B. L., Khosla, S. & Melton, L. J., III (2002) Endocr. Rev. 23, 279-302. - PubMed
-
- Kousteni, S., Chen, J. R., Bellido, T., Han, L., Ali, A. A., O'Brien, C. A., Plotkin, L., Fu, Q., Mancino, A. T., Wen, Y., et al. (2002) Science 298, 843-846. - PubMed
-
- Pfeilschifter, J., Koditz, R., Pfohl, M. & Schatz, H. (2002) Endocr. Rev. 23, 90-119. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
