

Review
doi: 10.1172/JCI19462.
Messenger RNA reprogramming by spliceosome-mediated RNA trans-splicing
Affiliations
- PMID: 12925685
- PMCID: PMC171395
- DOI: 10.1172/JCI19462
Item in Clipboard
Review
Messenger RNA reprogramming by spliceosome-mediated RNA trans-splicing
J Clin Invest.
2003 Aug.
Abstract
In the human genome, the majority of protein-encoding genes are interrupted by introns, which are removed from primary transcripts by a macromolecular enzyme known as the spliceosome. Spliceosomes can constitutively remove all the introns in a primary transcript to yield a fully spliced mRNA or alternatively splice primary transcripts leading to the production of many different mRNAs from one gene. This review examines how spliceosomes can recombine two primary transcripts in trans to reprogram messenger RNAs.
Figures
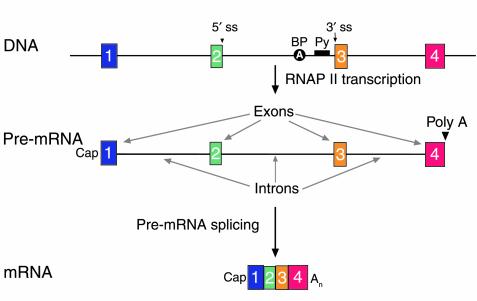
The structure of human genes and primary transcripts. A schematic of a gene (DNA), its primary transcript (pre-mRNA), and the mature mRNA product. The gene in this schematic spans four exons (boxes 1 through 4) and three introns. Genomic landmarks are indicated for the middle intron: the 5′ splice site (5′ ss), the branchpoint (BP) adenosine, the polypyrimidine tract (Py), and the 3′ splice site (3′ ss). The structure of the pre-mRNA, which is synthesized by RNA polymerase II, is shown. The 5′ end of the pre-mRNA is determined by transcription initiation and modified by a cap structure (27). The 3′ end is created and modified by cleavage and polyadenylation (poly A). Introns are removed in pre-mRNA splicing to yield an mRNA. On average, introns are longer than 3,000 nucleotides. The 5′ and 3′ terminal exons (1 and 4) average 300 nucleotides in length; internal exons (2 and 3) average 145 nucleotides in length (14, 15).
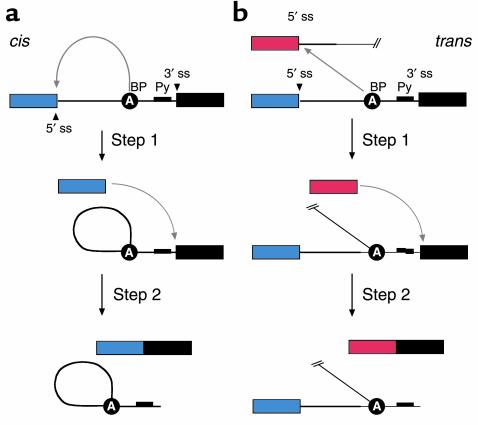
Both cis-splicing and trans-splicing reactions proceed via two phosphoryl transfer reactions. (a) The cis reaction. A schematic of the two phosphoryl transfer reactions required for intron removal (16). The exons are indicated as boxes (the first or 5′ exon is gray and the second or 3′ exon is black), and the intron is depicted as a line. In the first reaction (step 1), the 2′ OH group of a bulged adenosine at the branchpoint attacks the 5′ phosphate of the first residue of the intron forming the lariat intermediate and the “free” 5′ exon. The lariat contains the branched adenosine, so called because it is connected via conventional 5′ and 3′ links, but also contains a 2′-5′ linkage to the first residue of the intron (see ref. for a more extensive discussion of the splicing reactions). In the second step, the 3′ OH group of the last residue of the “free” 5′ exon attacks the 5′ phosphate of the first residue of the second exon, forming a product with the two exons ligated and releasing the intron as a lariat. (b) The trans reaction. A schematic of the two phosphoryl transfer reactions required for SMaRT. Icons are as described above, except that the trans-splicing or invading exon is shown as a red box. In step 1, the 2′ OH group of the bulged or branchpoint adenosine is again the nucleophile and is attacking the 5′ phosphate of the first residue of an intron in a second RNA molecule (this molecule could be identical in sequence to the first or it could be a completely different RNA). Because the reaction proceeds in trans the branched molecule is now a Y-shaped molecule, not a lariat. Step 2 proceeds as described for step 2 in the cis reaction; however, the exon product includes sequences from two RNAs.

Consensus sequences at and around the splice sites in nuclear pre-mRNA introns of mammals (17). Two types of introns, U2-dependent and U12-dependent, are spliced by two spliceosomes that share some components. The U2-dependent introns are by far the more prevalent. Although all the published SMaRT reactions to date have targeted U2-dependent introns, there is reason to believe that U12-dependent introns, which represent 0.1% of introns (14), will also be available for targeted SMaRT.
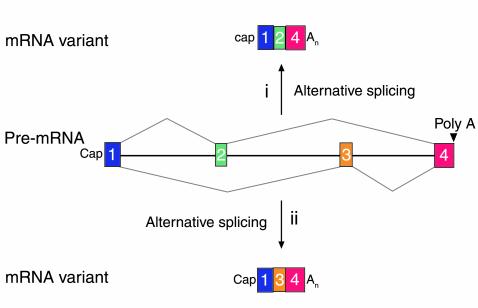
A schematic of two mRNA variants derived from alternative splicing of a primary transcript. In situation i, exon 2 is recognized whereas exon 3 is not and is read as being part of a large intron. In contrast, the reverse is true for situation ii. In many instances, alternative splicing is tightly regulated in cell type–specific fashion (27). An, refers to the polyA tail at the 3′ end of mRNAs.
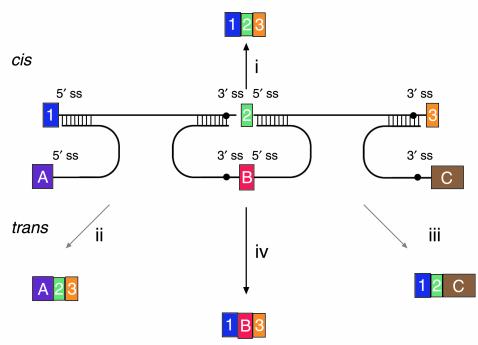
Versatility of mRNA reprogramming by targeted SMaRT. The figure shows a schematic of splicing reactions involving a three-exon pre-mRNA and PTMs. A conventional constitutive cis-splicing reaction leading to the production of the expected 1•2•3 mRNA is shown (i). Three targeted SMaRT reactions are shown. PTM[A] contains a functional 5′ splice site that can trans-splice to the 3′ splice site adjacent to exon 2 in the pre-mRNA target (ii). This trans-splicing produces a chimeric A•2•3 mRNA. PTM[A] is targeted to occlude the naturally occurring 5′ splice site at exon 1 to reduce the use of this site. PTM[C] contains a functional 3′ splice site that can trans-splice to the 5′ splice site adjacent to exon 2 in the pre-mRNA target (iii). This trans-splicing produces a chimeric 1•2•C mRNA. Finally, PTM[B] contains both 3′ and 5′ splice sites bordering an exon, and these splice sites can trans-splice with the 5′ splice site adjacent to exon 1 and the 5′ splice site adjacent to exon 3, respectively (iv). These two trans-splicing reactions lead to internal exon replacement and produce the chimeric 1•B•3 mRNA.
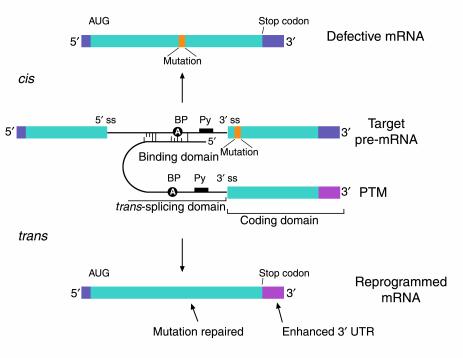
The anatomy of a PTM. The figure depicts a mutant target pre-mRNA (see the LacZ model system in ref. 12) that is incapable of coding for functional protein because it contains a mutation in the second exon (indicated as orange segment). The product of the conventional cis-splicing reaction for this pre-mRNA is a defective mRNA. The PTM shown contains several domains. The binding domain (also referred to as the targeting domain) binds to the pre-mRNA target, thus localizing the PTM near the site of the desired trans-splicing reaction. The binding domain can also be designed to occlude important elements within the 3′ splice site of the target and thus reduce the cis-splicing reaction. Between the binding domain and the trans-splicing domain there is usually a spacer region for flexibility. The trans-splicing domain, which in the case shown includes the elements required to make a potent 3′ splice site, is responsible for the reactivity of the PTM. Finally, the coding domain contains the necessary genetic information that will be imparted into the reprogrammed RNA. This can include protein-coding instructions as well as instructions for the effective processing, transport, and localization of the reprogrammed mRNA. In the example shown, the open reading frame has been changed to repair the mutation in the gene, and the 3′ untranslated region (3′ UTR) has been enhanced to increase mRNA stability and translation. It should be noted that the cis- and trans-splicing reactions are in competition. This implies that as the level of reprogrammed mRNA increases, the level of defective mRNA decreases. A decrease in the level of defective mRNA may be very useful in the case of dominant mutations.
References
-
- Kole R, Sazani P. Antisense effects in the nucleus: modification of splicing. Curr. Opin. Mol. Ther. 2001;3:229–234. - PubMed
-
- Sullenger BA, Gilboa E. Emerging clinical applications of RNA. Nature. 2002;418:252–258. - PubMed
-
- Garcia-Blanco MA, Puttaraju M, Mansfield SG, Mitchell LG. Spliceosome-mediated RNA trans-splicing in gene therapy and genomics. Gene Ther. Reg. 2000;1:141–163. - PubMed
-
- Sullenger B, Cech TR. Ribozyme-mediated repair of defective mRNA by targeted trans-splicing. Nature. 1994;317:619–622. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
