

Role for integrin-linked kinase in mediating tubular epithelial to mesenchymal transition and renal interstitial fibrogenesis
- PMID: 12925691
- PMCID: PMC171389
- DOI: 10.1172/JCI17913
Role for integrin-linked kinase in mediating tubular epithelial to mesenchymal transition and renal interstitial fibrogenesis
Erratum in
- J Clin Invest. 2004 Feb;113(3):491
Abstract
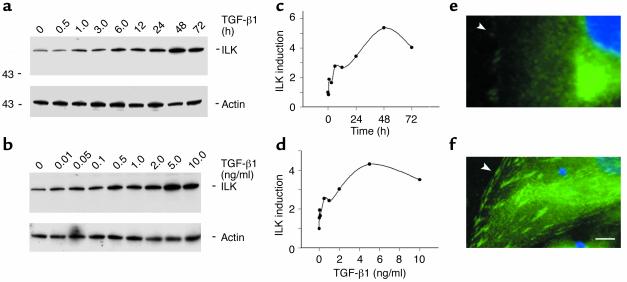
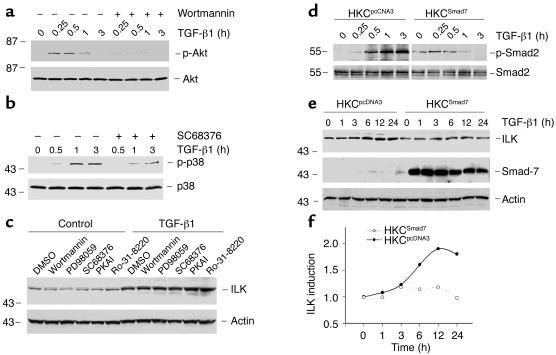
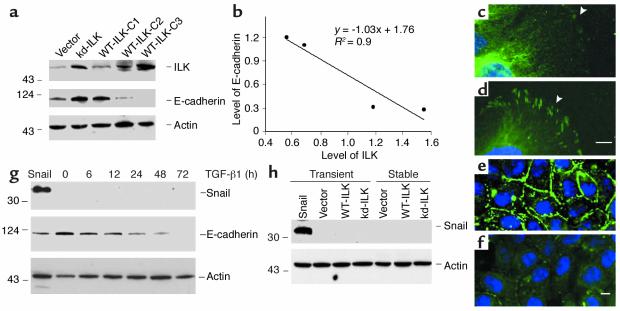
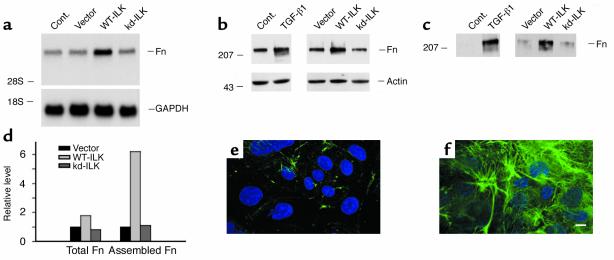
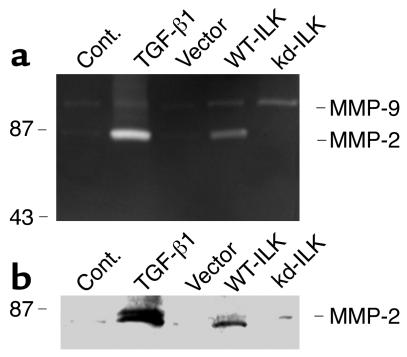
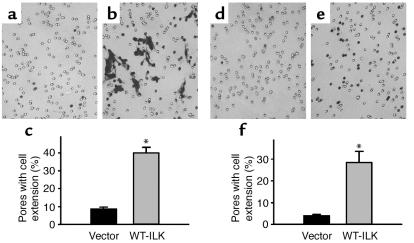
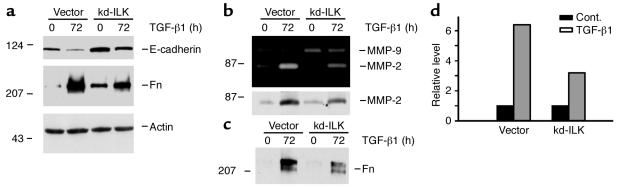
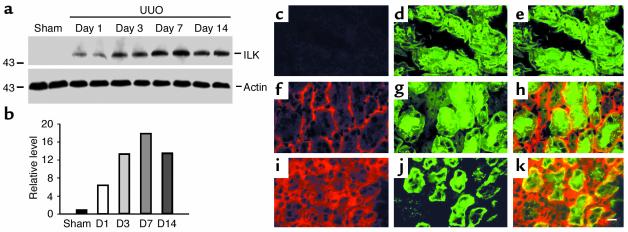
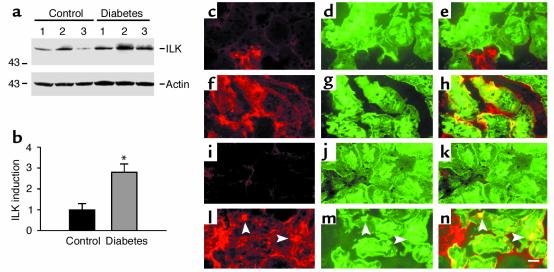
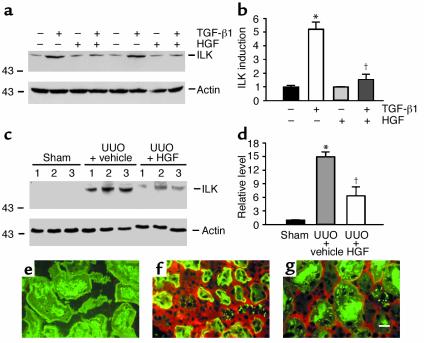
Under pathologic conditions, renal tubular epithelial cells can undergo epithelial to mesenchymal transition (EMT), a phenotypic conversion that is believed to play a critical role in renal interstitial fibrogenesis. However, the underlying mechanism that governs this process remains largely unknown. Here we demonstrate that integrin-linked kinase (ILK) plays an important role in mediating tubular EMT induced by TGF-beta1. TGF-beta1 induced ILK expression in renal tubular epithelial cells in a time- and dose-dependent manner, which was dependent on intracellular Smad signaling. Forced expression of ILK in human kidney proximal tubular epithelial cells suppressed E-cadherin expression and induced fibronectin expression and its extracellular assembly. ILK also induced MMP-2 expression and promoted cell migration and invasion in Matrigel. Conversely, ectopic expression of a dominant-negative, kinase-dead form of ILK largely abrogated TGF-beta1-initiated tubular cell phenotypic conversion. In vivo, ILK was markedly induced in renal tubular epithelia in mouse models of chronic renal diseases, and such induction was spatially and temporally correlated with tubular EMT. Moreover, inhibition of ILK expression by HGF was associated with blockade of tubular EMT and attenuation of renal fibrosis. These findings suggest that ILK is a critical mediator for tubular EMT and likely plays a crucial role in the pathogenesis of chronic renal fibrosis.
Figures
Similar articles
-
PINCH-1 promotes tubular epithelial-to-mesenchymal transition by interacting with integrin-linked kinase.J Am Soc Nephrol. 2007 Sep;18(9):2534-43. doi: 10.1681/ASN.2007030315. Epub 2007 Jul 26. J Am Soc Nephrol. 2007. PMID: 17656471
-
A novel mechanism by which hepatocyte growth factor blocks tubular epithelial to mesenchymal transition.J Am Soc Nephrol. 2005 Jan;16(1):68-78. doi: 10.1681/ASN.2003090795. Epub 2004 Nov 10. J Am Soc Nephrol. 2005. PMID: 15537870
-
Inhibition of integrin-linked kinase attenuates renal interstitial fibrosis.J Am Soc Nephrol. 2009 Sep;20(9):1907-18. doi: 10.1681/ASN.2008090930. Epub 2009 Jun 18. J Am Soc Nephrol. 2009. PMID: 19541809 Free PMC article.
-
The role of tubular epithelial-mesenchymal transition in progressive kidney disease.Cells Tissues Organs. 2007;185(1-3):222-31. doi: 10.1159/000101323. Cells Tissues Organs. 2007. PMID: 17587828 Review.
-
The role of epithelial-to-mesenchymal transition in renal fibrosis.J Mol Med (Berl). 2004 Mar;82(3):175-81. doi: 10.1007/s00109-003-0517-9. Epub 2004 Jan 30. J Mol Med (Berl). 2004. PMID: 14752606 Review.
Cited by
-
Lefty A attenuates the TGF-beta1-induced epithelial to mesenchymal transition of human renal proximal epithelial tubular cells.Mol Cell Biochem. 2010 Jun;339(1-2):263-70. doi: 10.1007/s11010-010-0389-6. Epub 2010 Feb 16. Mol Cell Biochem. 2010. PMID: 20157767
-
Integrin-linked kinase (ILK): the known vs. the unknown and perspectives.Cell Mol Life Sci. 2022 Jan 28;79(2):100. doi: 10.1007/s00018-021-04104-1. Cell Mol Life Sci. 2022. PMID: 35089438 Free PMC article. Review.
-
Hepatocyte growth factor inhibits epithelial to myofibroblast transition in lung cells via Smad7.Am J Respir Cell Mol Biol. 2009 Jun;40(6):643-53. doi: 10.1165/rcmb.2008-0217OC. Epub 2008 Nov 6. Am J Respir Cell Mol Biol. 2009. PMID: 18988920 Free PMC article.
-
Lack of α8 integrin leads to morphological changes in renal mesangial cells, but not in vascular smooth muscle cells.BMC Cell Biol. 2010 Dec 31;11:102. doi: 10.1186/1471-2121-11-102. BMC Cell Biol. 2010. PMID: 21194485 Free PMC article.
-
Effect of Huayu Tongluo Herbs on Reduction of Proteinuria via Inhibition of Wnt/β-Catenin Signaling Pathway in Diabetic Rats.Evid Based Complement Alternat Med. 2017;2017:3054145. doi: 10.1155/2017/3054145. Epub 2017 Jun 1. Evid Based Complement Alternat Med. 2017. PMID: 28656052 Free PMC article.
References
-
- Strutz F, Muller GA. Transdifferentiation comes of age. Nephrol. Dial. Transplant. 2000;15:1729–1731. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
