

Maintenance of stem cell populations in plants
- PMID: 12930889
- PMCID: PMC304093
- DOI: 10.1073/pnas.1834206100
Maintenance of stem cell populations in plants
Abstract
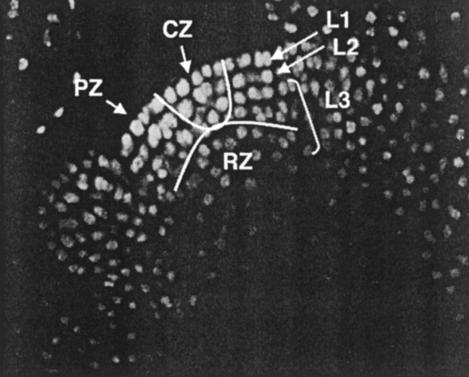
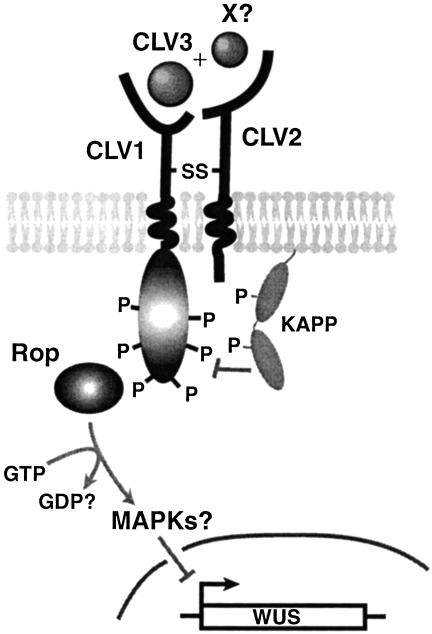
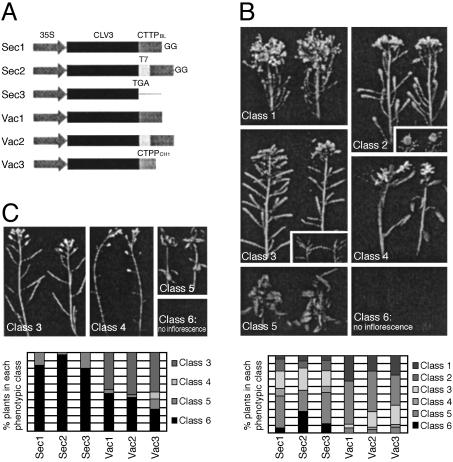
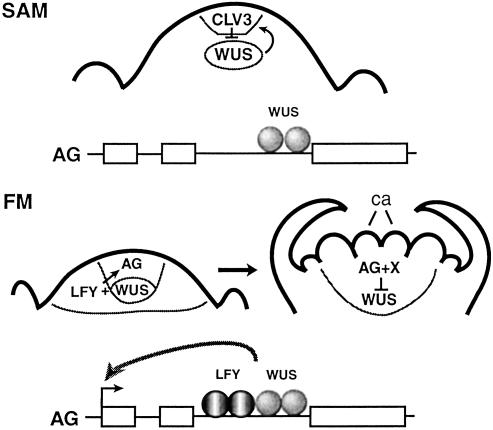
Flowering plants have the unique ability to produce new organs continuously, for hundreds of years in some species, from stem cell populations maintained at their actively growing tips. The shoot tip is called the shoot apical meristem, and it acts as a self-renewing source of undifferentiated, pluripotent stem cells whose descendents become incorporated into organ and tissue primordia and acquire different fates. Stem cell maintenance is an active process, requiring constant communication between different regions of the shoot apical meristem to coordinate loss of stem cells from the meristem through differentiation with their replacement through cell division. Stem cell research in model plant systems is facilitated by the fact that mutants with altered meristem cell identity or accumulation are viable, allowing dissection of stem cell behavior by using genetic, molecular, and biochemical methods. Such studies have determined that in the model plant Arabidopsis thaliana stem cell maintenance information flows via a signal transduction pathway that is established during embryogenesis and maintained throughout the life cycle. Signaling through this pathway results in the generation of a spatial feedback loop, involving both positive and negative interactions, that maintains stem cell homeostasis. Stem cell activity during reproductive development is terminated by a temporal feedback loop involving both stem cell maintenance genes and a phase-specific flower patterning gene. Our current investigations provide additional insights into the molecular mechanisms that regulate stem cell activity in higher plants.
Figures
References
-
- Meyerowitz, E. M. (1997) Cell 88, 299-308. - PubMed
-
- Long, J. A. & Barton, M. K. (1998) Development (Cambridge, U.K.) 125, 3027-3035. - PubMed
-
- Steeves, T. A. & Sussex, I. M. (1989) Patterns in Plant Development (Cambridge Univ. Press, New York).
-
- Lyndon, R. F. (1998) The Shoot Apical Meristem: Its Growth and Development (Cambridge Univ. Press, Cambridge, U.K.).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
