

Analysis of the DNA joining repertoire of Chlorella virus DNA ligase and a new crystal structure of the ligase-adenylate intermediate
- PMID: 12930960
- PMCID: PMC212790
- DOI: 10.1093/nar/gkg665
Analysis of the DNA joining repertoire of Chlorella virus DNA ligase and a new crystal structure of the ligase-adenylate intermediate
Abstract
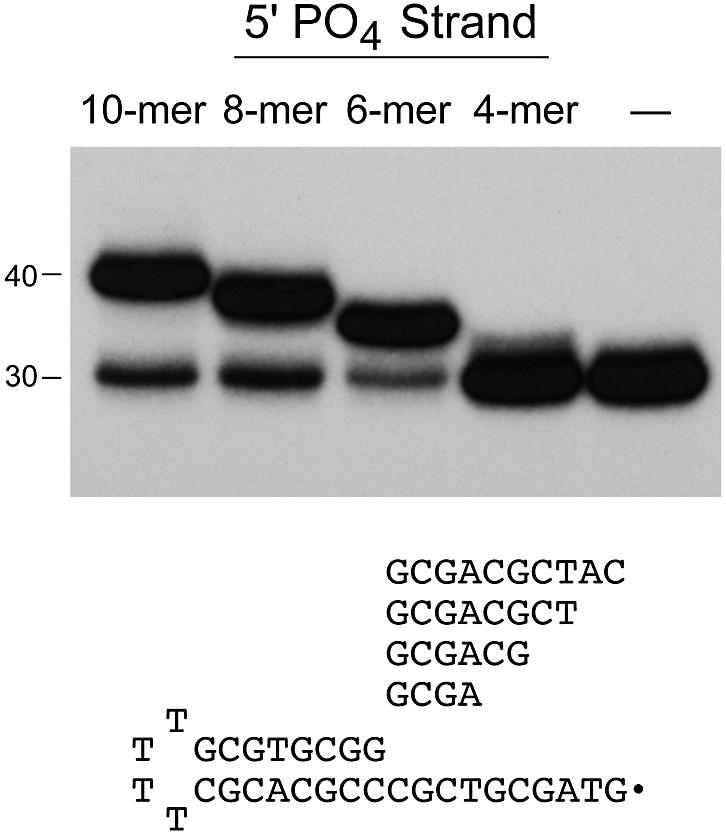
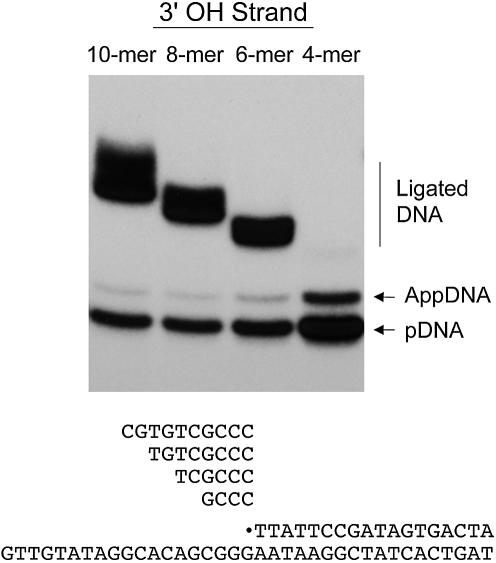
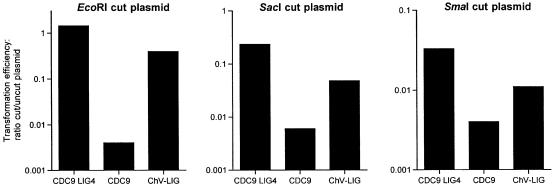
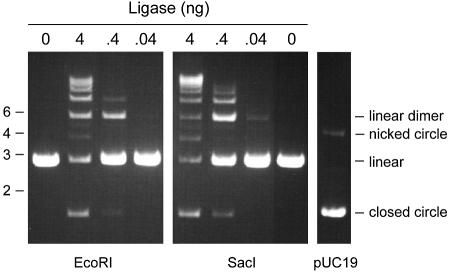
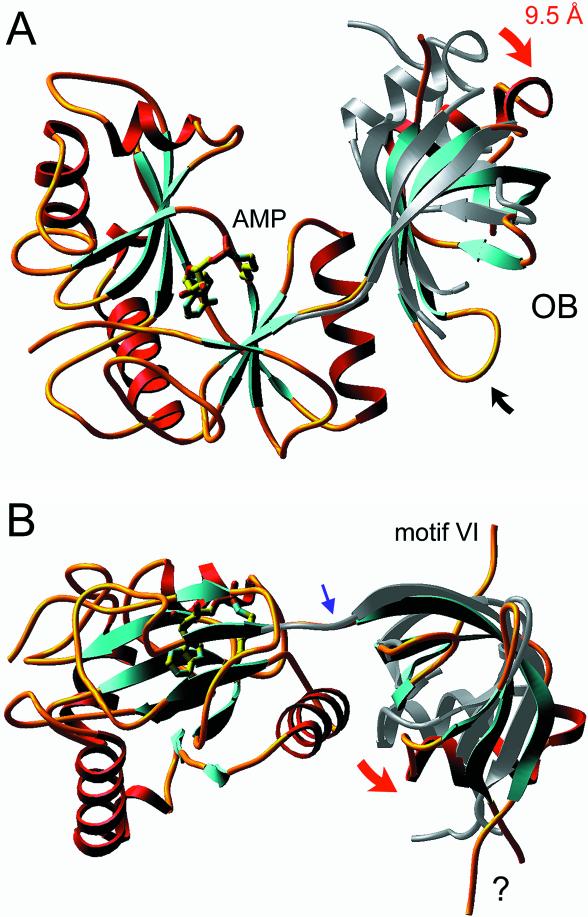
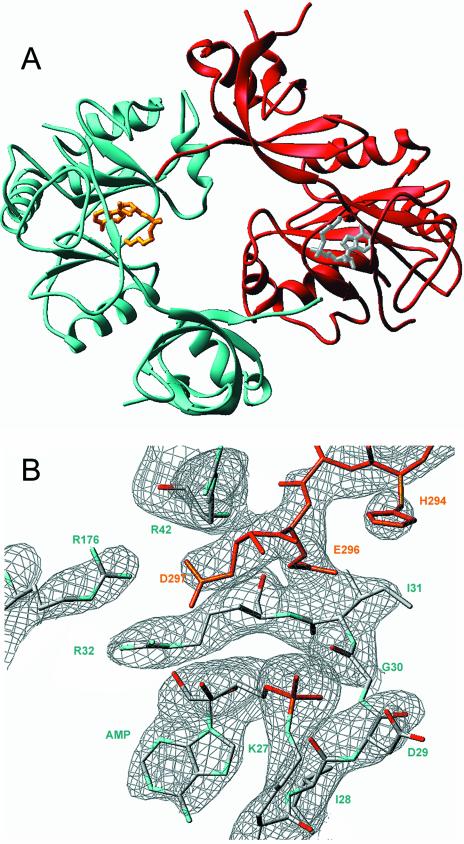
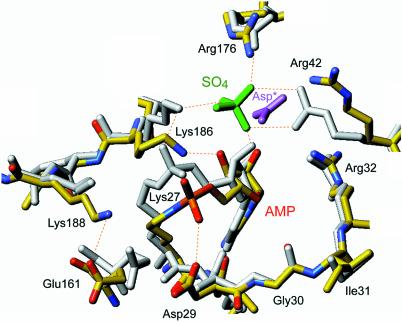
Chlorella virus DNA ligase is the smallest eukaryotic ATP-dependent DNA ligase known; it suffices for yeast cell growth in lieu of the essential yeast DNA ligase Cdc9. The Chlorella virus ligase-adenylate intermediate has an intrinsic nick sensing function and its DNA footprint extends 8-9 nt on the 3'-hydroxyl (3'-OH) side of the nick and 11-12 nt on the 5'-phosphate (5'-PO4) side. Here we establish the minimal length requirements for ligatable 3'-OH and 5'-PO4 strands at the nick (6 nt) and describe a new crystal structure of the ligase-adenylate in a state construed to reflect the configuration of the active site prior to nick recognition. Comparison with a previous structure of the ligase-adenylate bound to sulfate (a mimetic of the nick 5'-PO4) suggests how the positions and contacts of the active site components and the bound adenylate are remodeled by DNA binding. We find that the minimal Chlorella virus ligase is capable of catalyzing non-homologous end-joining reactions in vivo in yeast, a process normally executed by the structurally more complex cellular Lig4 enzyme. Our results suggest a model of ligase evolution in which: (i) a small 'pluripotent' ligase is the progenitor of the much larger ligases found presently in eukaryotic cells and (ii) gene duplications, variations within the core ligase structure and the fusion of new domains to the core structure (affording new protein-protein interactions) led to the compartmentalization of eukaryotic ligase function, i.e. by enhancing some components of the functional repertoire of the ancestral ligase while disabling others.
Figures
References
-
- Weiss B., Thompson,A. and Richardson,C.C. (1968) Enzymatic breakage and rejoining of deoxyribonucleic acid: properties of the enzyme–adenylate intermediate in the polynucleotide ligase reaction. J. Biol. Chem., 246, 4523–4530. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
