

Inhibition of proteasomal activity causes inclusion formation in neuronal and non-neuronal cells overexpressing Parkin
- PMID: 12937272
- PMCID: PMC266771
- DOI: 10.1091/mbc.e03-02-0078
Inhibition of proteasomal activity causes inclusion formation in neuronal and non-neuronal cells overexpressing Parkin
Abstract
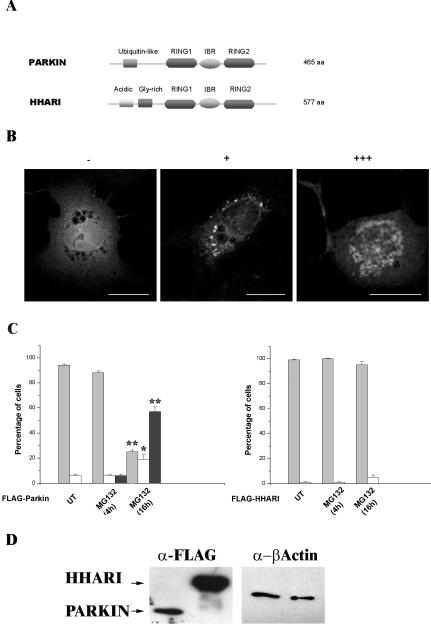
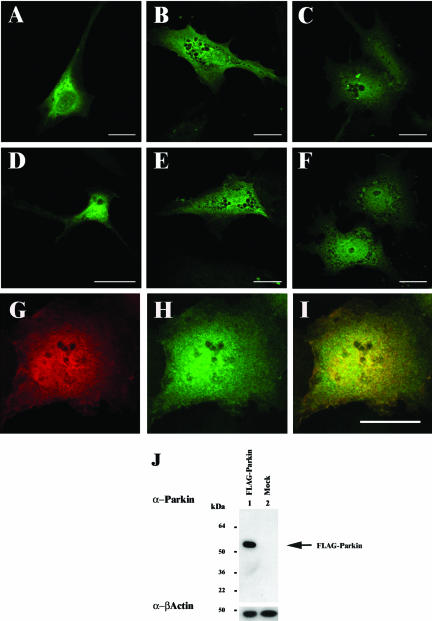
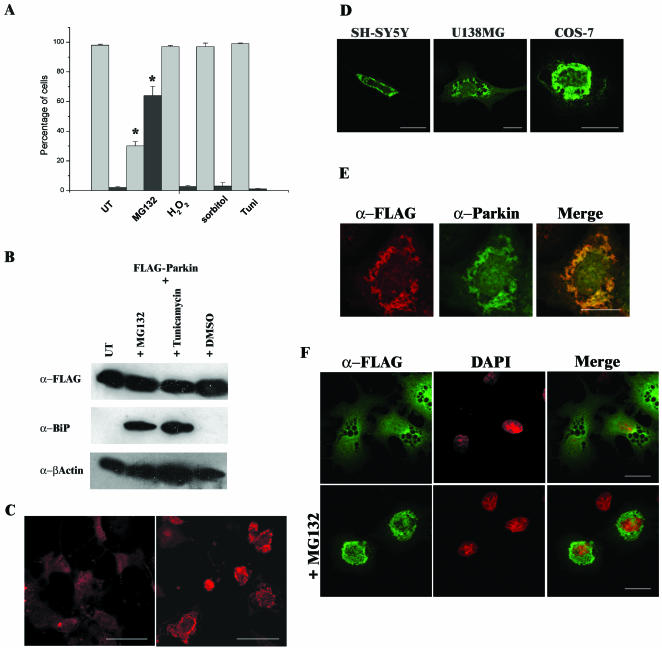
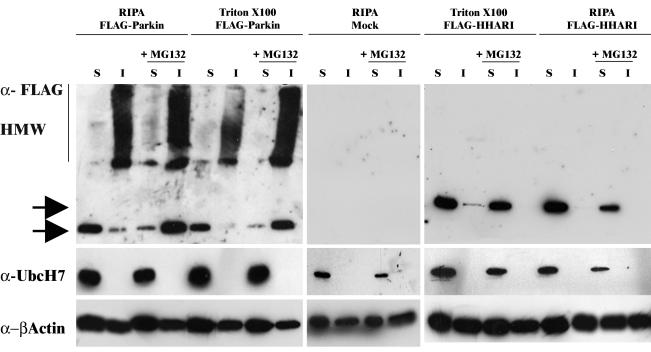
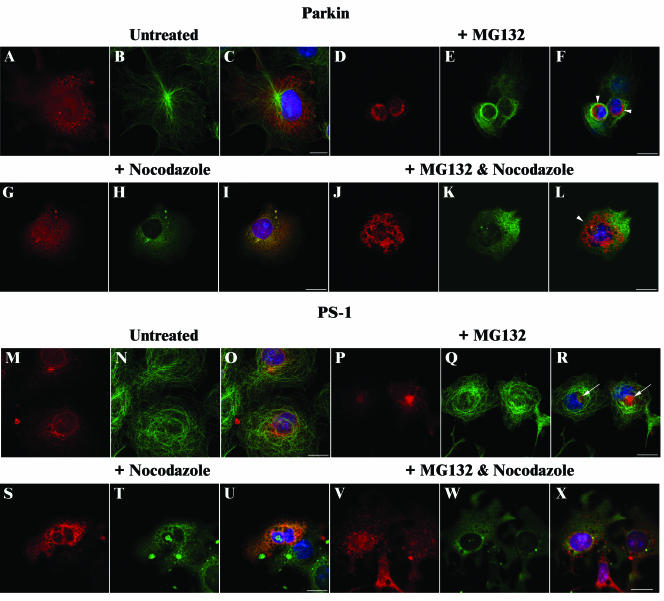
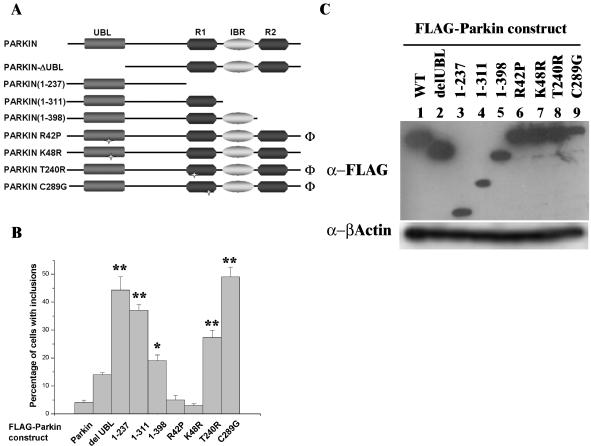
Association between protein inclusions and neurodegenerative diseases, including Parkinson's and Alzheimer's diseases, and polyglutamine disorders, has been widely documented. Although ubiquitin is conjugated to many of these aggregated proteins, the 26S proteasome does not efficiently degrade them. Mutations in the ubiquitin-protein ligase Parkin are associated with autosomal recessive juvenile Parkinsonism. Although Parkin-positive inclusions are not detected in brains of autosomal recessive juvenile Parkinsonism patients, Parkin is found in Lewy bodies in sporadic disease. This suggests that loss of Parkin ligase activity via mutation, or sequestration to Lewy bodies, is a contributory factor to sporadic disease onset. We now demonstrate that decreased proteasomal activity causes formation of large, noncytotoxic inclusions within the cytoplasm of both neuronal and nonneuronal cells overexpressing Parkin. This is not a general phenomenon as there is an absence of similar inclusions when HHARI, a structural homolog of Parkin, is overexpressed. The inclusions colocalize with ubiquitin and with proteasomes. Furthermore, Parkin inclusions colocalize with gamma-tubulin, acetylated alpha-tubulin, and cause redistribution of vimentin, suggesting aggresome-like properties. Our data imply that lower proteasomal activity, previously observed in brain tissue of Parkinson's disease patients, leads to Parkin accumulation and a concomitant reduction in ligase activity, thereby promoting Lewy body formation.
Figures


References
-
- Ardley, H.C., Tan, N.G.S., Rose, S.A., Markham, A.F., and Robinson, P.A. (2001). Features of the Parkin/Ariadne-like ubiquitin ligase, HHARI, that regulate its interaction with the ubiquitin-conjugating enzyme, UbcH7. J. Biol. Chem. 276, 19640-19647. - PubMed
-
- Arima, K., Mizutani, T., Alim, M.A., Tonozuka-Uehara, H., Izumiyama, Y., Hirai, S., and Ueda, K. (2000). NACP/alpha-synuclein and tau constitute two distinctive subsets of filaments in the same neuronal inclusions in brains from a family of parkinsonism and dementia with Lewy bodies: double-immunolabeling fluorescence and electron microscopic studies. Acta Neuropathol. 100, 115-121. - PubMed
-
- Bence, N.F., Sampat, R.M., and Kopito, R.R. (2001). Impairment of the ubiquitin-proteasome system by protein aggregation. Science 292, 1552-1555. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
