

DNA damage modulates nucleolar interaction of the Werner protein with the AAA ATPase p97/VCP
- PMID: 12937274
- PMCID: PMC207013
- DOI: 10.1091/mbc.e03-02-0111
DNA damage modulates nucleolar interaction of the Werner protein with the AAA ATPase p97/VCP
Abstract
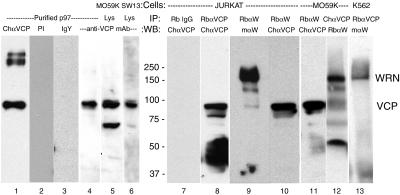
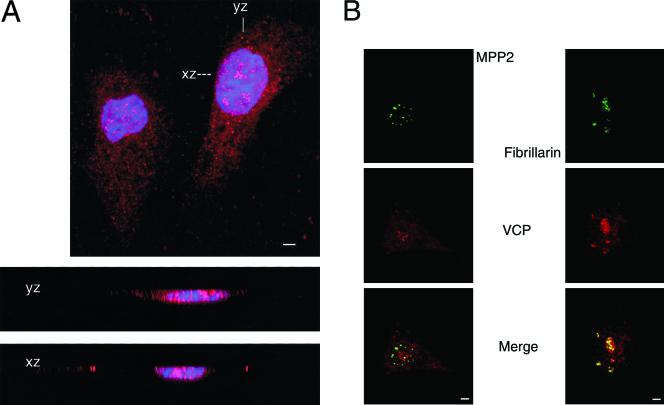
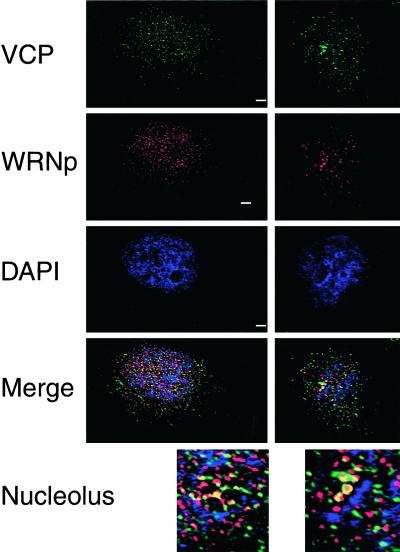
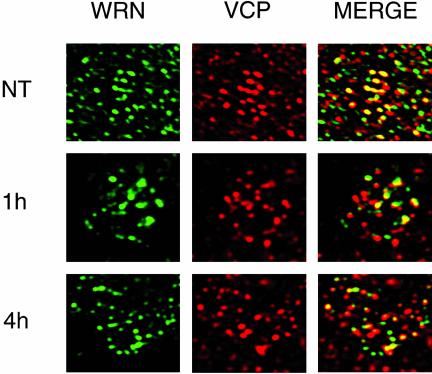
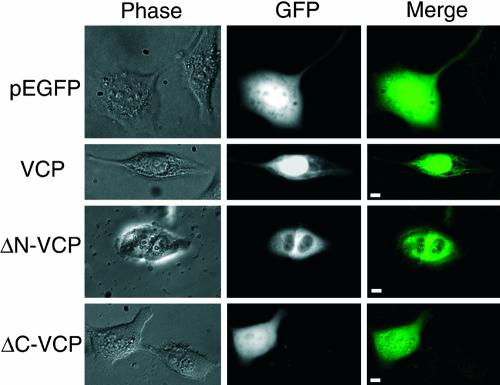
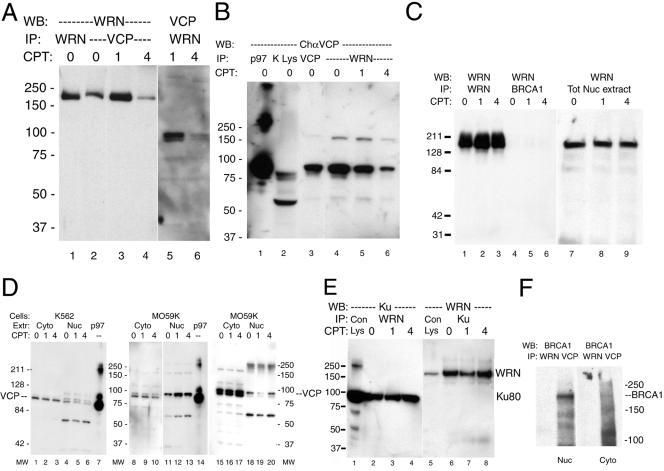
We report a novel nucleolar interaction between the AAA ATPase p97/VCP and the Werner protein (WRNp), a member of the RecQ helicase family. p97/VCP mediates several important cellular functions in eucaryotic cells, including membrane fusion of the endoplasmic reticulum and Golgi and ubiquitin-dependent protein degradation. Mutations in the WRN gene cause Werner syndrome, a genetic disorder characterized by premature onset of aging symptoms, a higher incidence of cancer, and a high susceptibility to DNA damage caused by topoisomerase inhibitors. We observed that both WRNp and valosin-containing protein (VCP) were present in the nucleoplasm and in nucleolar foci in mammalian cells and that WRNp and p97/VCP physically interacted in the nucleoli. Importantly, the nucleolar WRNp/VCP complex was dissociated by treatment with camptothecin, an inhibitor of topoisomerase I, whereas other WRNp-associated protein complexes, such as WRNp/Ku 80, were not dissociated by this drug. Because WRN syndrome cells are sensitive to topoisomerase inhibitors, these observations suggest that the VCP/WRNp interaction plays an important role in WRN biology. We propose a novel role for VCP in the DNA damage response pathway through modulation of WRNp availability.
Figures
References
-
- Abmayr, S.M., and Workman, J.L. (2001). Unit 12.1 Preparation of nuclear and cytoplasmic extracts from mammalian cells. In: Current Protocols in Molecular Biology Online, ed. F.M. Ausubel, New York: John Wiley & Sons. - PubMed
-
- Acharya, U., Jacobs, R., Peters, J.M., Watson, N., Farquhar, M.G., and Malhotra, V. (1995). The formation of Golgi stacks from vesiculated Golgi membranes requires two distinct fusion events. Cell 82, 895–904. - PubMed
-
- Andersen, J.S., Lyon, C.E., Fox, A.H., Leung, A.K., Lam, Y.W., Steen, H., Mann, M., and Lamond, A.I. (2002). Directed proteomic analysis of the human nucleolus. Curr. Biol. 12, 1–11. - PubMed
-
- Blander, G., Kipnis, J., Leal, J.F., Yu, C.E., Schellenberg, G.D., and Oren, M. (1999). Physical and functional interaction between p53 and the Werner's syndrome protein. J. Biol. Chem. 274, 29463–29469. - PubMed
-
- Blander, G., Zalle, N., Leal, J.F., Bar-Or, R.L., Yu, C.E., and Oren, M. (2000). The Werner syndrome protein contributes to induction of p53 by DNA damage. FASEB J. 14, 2138–2140. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
