

A naturally selected dimorphism within the HLA-B44 supertype alters class I structure, peptide repertoire, and T cell recognition
- PMID: 12939341
- PMCID: PMC2194191
- DOI: 10.1084/jem.20030066
A naturally selected dimorphism within the HLA-B44 supertype alters class I structure, peptide repertoire, and T cell recognition
Abstract
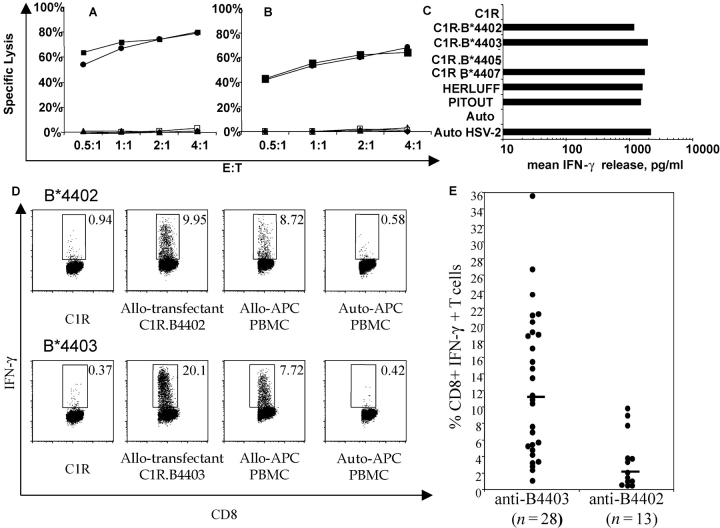
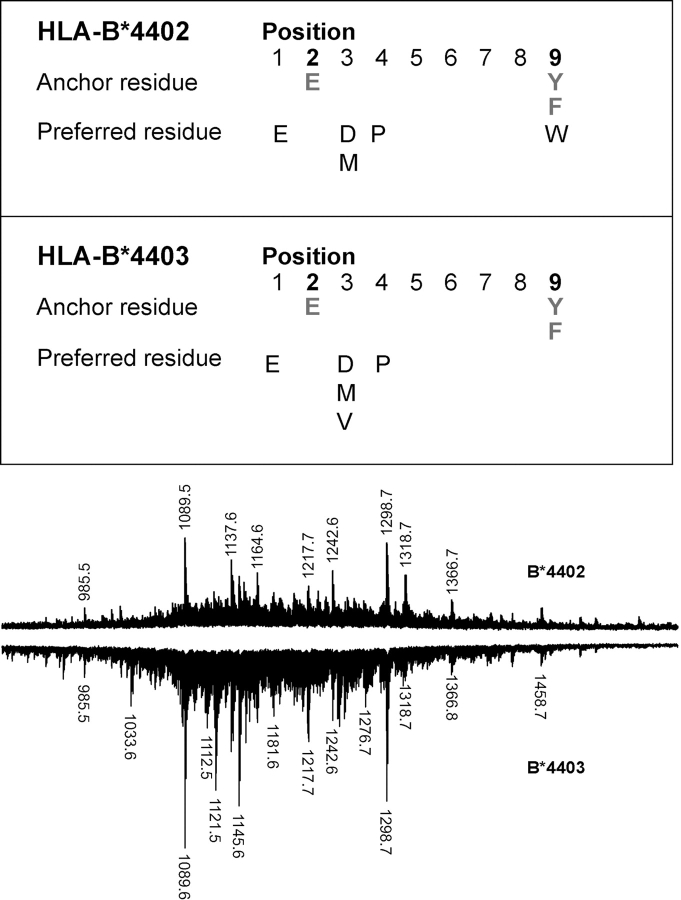
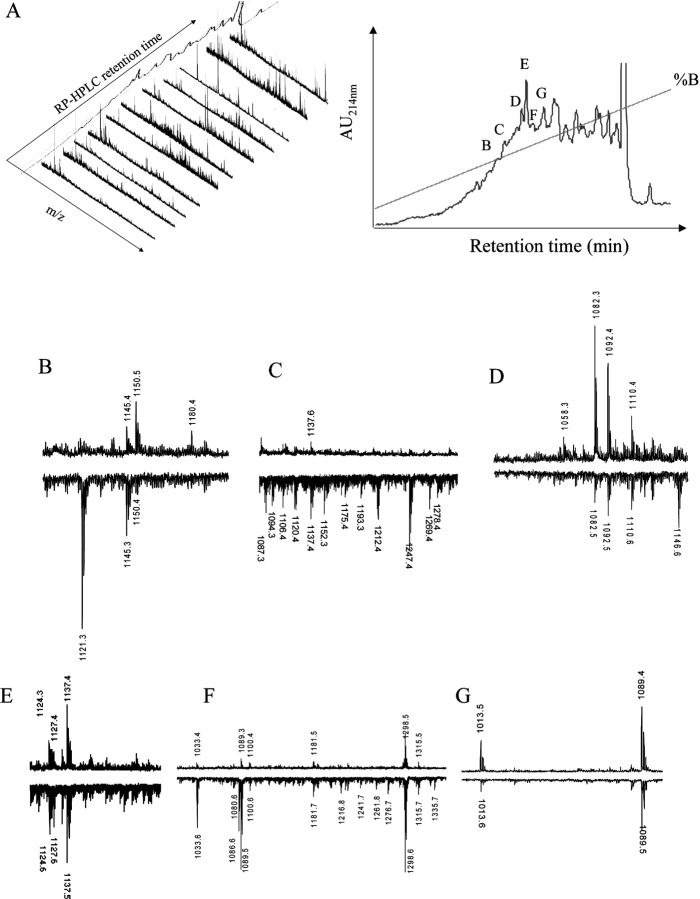
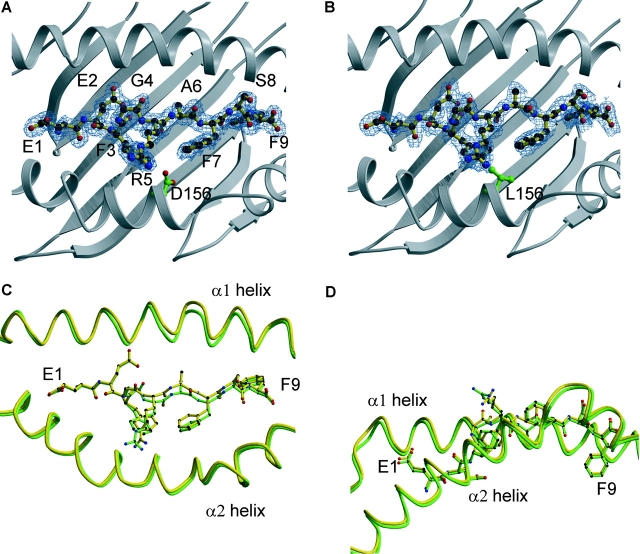
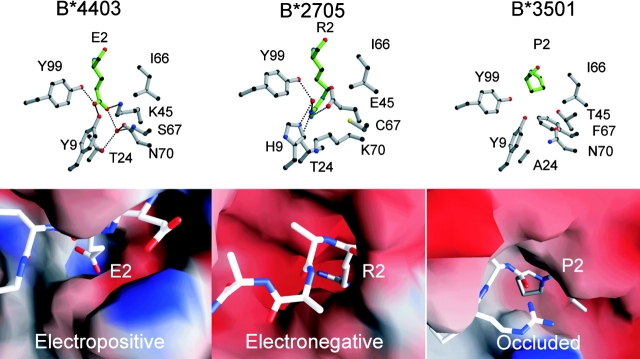
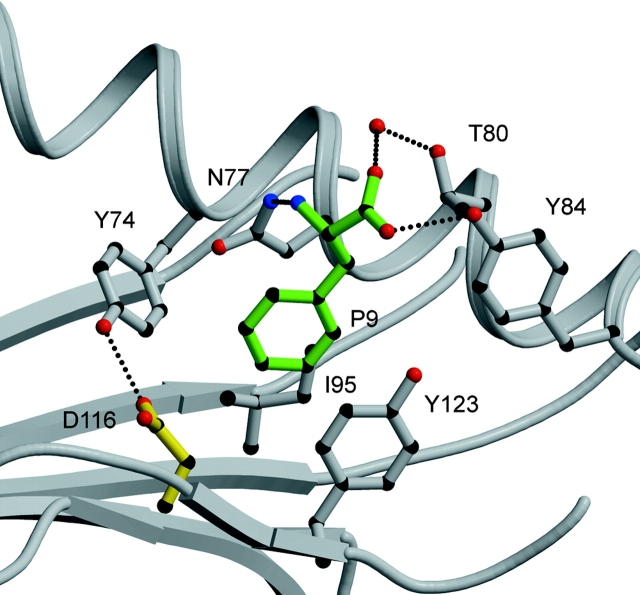
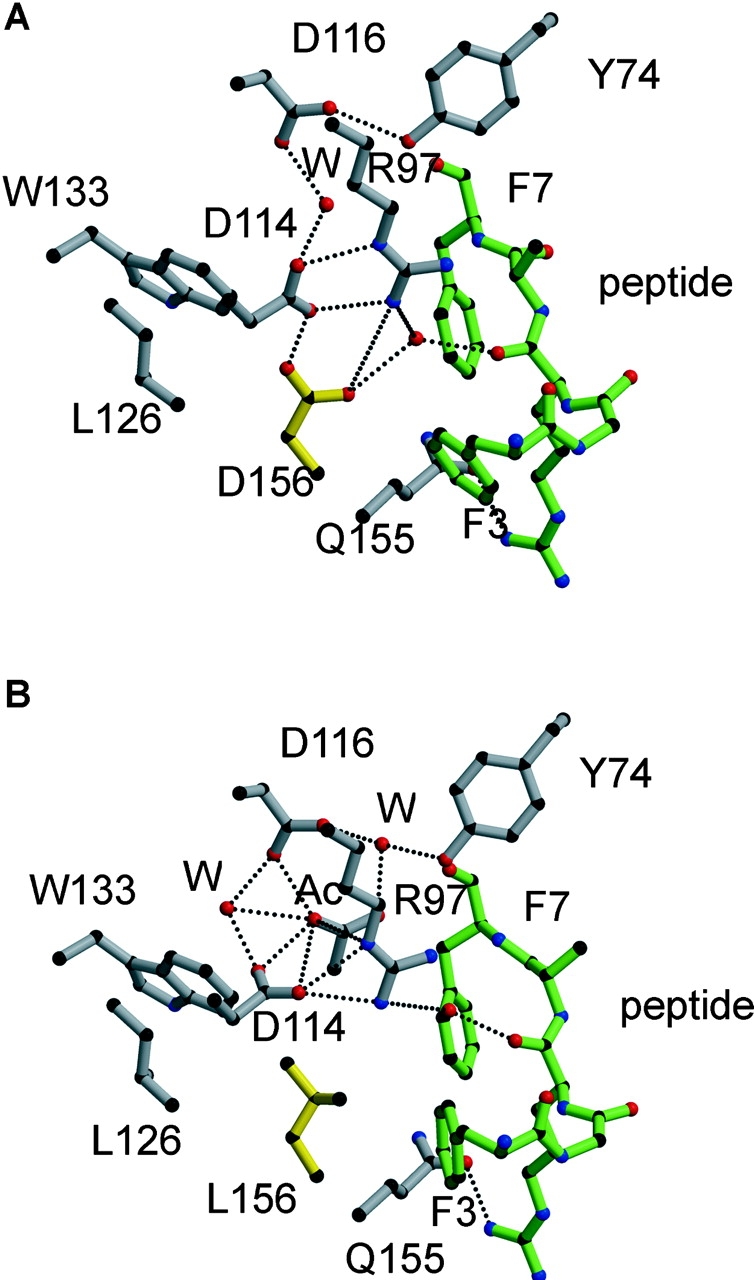
HLA-B*4402 and B*4403 are naturally occurring MHC class I alleles that are both found at a high frequency in all human populations, and yet they only differ by one residue on the alpha2 helix (B*4402 Asp156-->B*4403 Leu156). CTLs discriminate between HLA-B*4402 and B*4403, and these allotypes stimulate strong mutual allogeneic responses reflecting their known barrier to hemopoeitic stem cell transplantation. Although HLA-B*4402 and B*4403 share >95% of their peptide repertoire, B*4403 presents more unique peptides than B*4402, consistent with the stronger T cell alloreactivity observed toward B*4403 compared with B*4402. Crystal structures of B*4402 and B*4403 show how the polymorphism at position 156 is completely buried and yet alters both the peptide and the heavy chain conformation, relaxing ligand selection by B*4403 compared with B*4402. Thus, the polymorphism between HLA-B*4402 and B*4403 modifies both peptide repertoire and T cell recognition, and is reflected in the paradoxically powerful alloreactivity that occurs across this "minimal" mismatch. The findings suggest that these closely related class I genes are maintained in diverse human populations through their differential impact on the selection of peptide ligands and the T cell repertoire.
Figures
References
-
- Hill, A.V., C.E. Allsopp, D. Kwiatkowski, N.M. Anstey, P. Twumasi, P.A. Rowe, S. Bennett, D. Brewster, A.J. McMichael, and B.M. Greenwood. 1991. Common west African HLA antigens are associated with protection from severe malaria. Nature. 352:595–600. - PubMed
-
- Parham, P., and T. Ohta. 1996. Population biology of antigen presentation by MHC class I molecules. Science. 272:67–74. - PubMed
-
- Messaoudi, I., J.A. Patino, R. Dyall, J. LeMaoult, and J. Nikolich-Zugich. 2002. Direct link between MHC polymorphism, T cell avidity, and diversity in immune defense. Science. 298:1797–1800. - PubMed
-
- Marsh, S.G., P. Parham, and L.D. Barber. 2000. The HLA Facts Book. Academic Press, London. 398 pp.
-
- Sette, A., and J. Sidney. 1999. Nine major HLA class I supertypes account for the vast preponderance of HLA-A and -B polymorphism. Immunogenetics. 50:201–212. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
