

FWD1-mediated degradation of FREQUENCY in Neurospora establishes a conserved mechanism for circadian clock regulation
- PMID: 12941694
- PMCID: PMC202367
- DOI: 10.1093/emboj/cdg425
FWD1-mediated degradation of FREQUENCY in Neurospora establishes a conserved mechanism for circadian clock regulation
Abstract
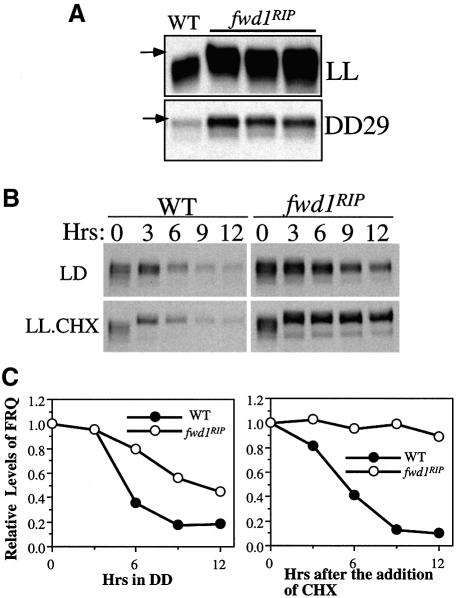
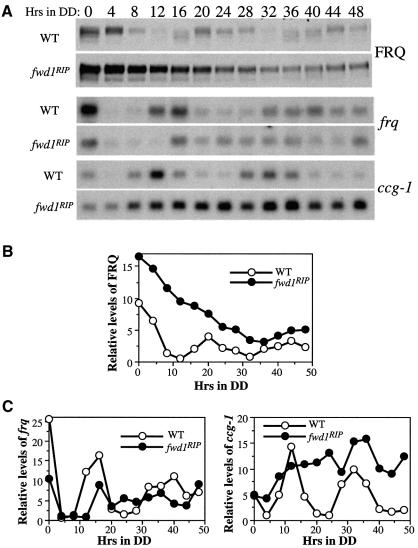
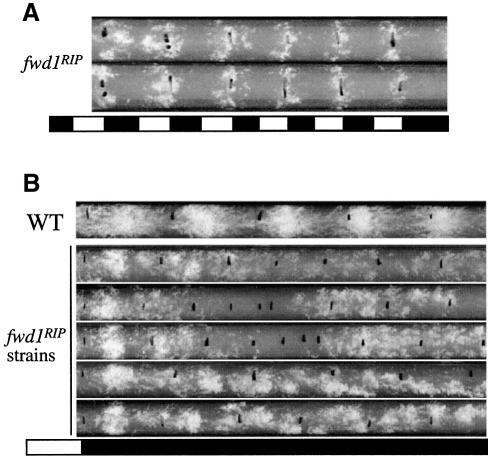
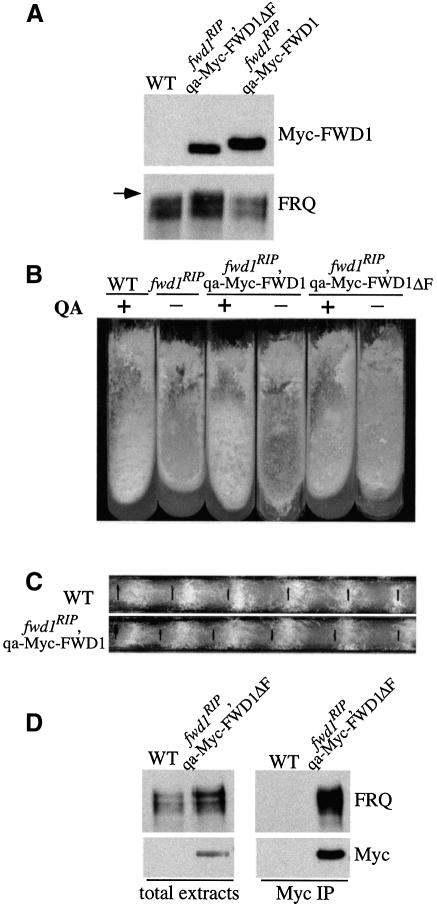
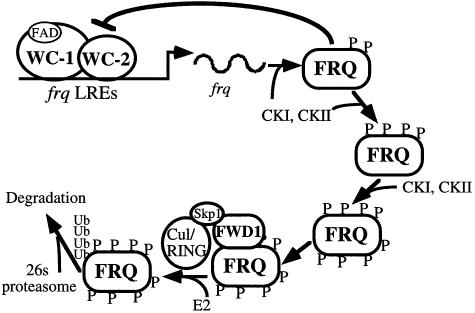
Phosphorylation of the Neurospora circadian clock protein FREQUENCY (FRQ) regulates its degradation and the proper function of the clock. The mechanism by which FRQ undergoes degradation has not been established. Here we show that FRQ is likely ubiquitylated in vivo, and its proper degradation requires FWD1, an F-box/WD-40 repeat-containing protein. In the fwd1 disruption strains, FRQ degradation is severely impaired, resulting in the accumulation of hyperphosphorylated FRQ. Furthermore, the circadian rhythms of gene expression and the circadian conidiation rhythms are abolished in these fwd1 mutants. Finally, FRQ and FWD1 interact physically in vivo, suggesting that FWD1 is the substrate-recruiting subunit of an SCF-type ubiquitin ligase responsible for FRQ ubiquitylation and degradation. Together with the recent finding that Slimb (the Drosophila homolog of FWD1) is involved in the degradation of the Period protein in flies, our results indicate that FWD1 regulates the degradation of FRQ in Neurospora and is an evolutionarily conserved component of the eukaryotic circadian clock.
Figures

Similar articles
-
Degradation of the Neurospora circadian clock protein FREQUENCY through the ubiquitin-proteasome pathway.Biochem Soc Trans. 2005 Nov;33(Pt 5):953-6. doi: 10.1042/BST20050953. Biochem Soc Trans. 2005. PMID: 16246019
-
Regulation of the Neurospora circadian clock by casein kinase II.Genes Dev. 2002 Apr 15;16(8):994-1006. doi: 10.1101/gad.965102. Genes Dev. 2002. PMID: 11959847 Free PMC article.
-
Phosphorylation of the Neurospora clock protein FREQUENCY determines its degradation rate and strongly influences the period length of the circadian clock.Proc Natl Acad Sci U S A. 2000 Jan 4;97(1):234-9. doi: 10.1073/pnas.97.1.234. Proc Natl Acad Sci U S A. 2000. PMID: 10618401 Free PMC article.
-
Posttranslational regulation of Neurospora circadian clock by CK1a-dependent phosphorylation.Cold Spring Harb Symp Quant Biol. 2007;72:177-83. doi: 10.1101/sqb.2007.72.025. Cold Spring Harb Symp Quant Biol. 2007. PMID: 18419275 Review.
-
A circadian clock in Neurospora: how genes and proteins cooperate to produce a sustained, entrainable, and compensated biological oscillator with a period of about a day.Cold Spring Harb Symp Quant Biol. 2007;72:57-68. doi: 10.1101/sqb.2007.72.072. Cold Spring Harb Symp Quant Biol. 2007. PMID: 18522516 Free PMC article. Review.
Cited by
-
Molecular mechanism of suppression of circadian rhythms by a critical stimulus.EMBO J. 2006 Nov 15;25(22):5349-57. doi: 10.1038/sj.emboj.7601397. Epub 2006 Oct 26. EMBO J. 2006. PMID: 17066078 Free PMC article.
-
Transcription factor CBF-1 is critical for circadian gene expression by modulating WHITE COLLAR complex recruitment to the frq locus.PLoS Genet. 2018 Sep 12;14(9):e1007570. doi: 10.1371/journal.pgen.1007570. eCollection 2018 Sep. PLoS Genet. 2018. PMID: 30208021 Free PMC article.
-
Protein kinase A and casein kinases mediate sequential phosphorylation events in the circadian negative feedback loop.Genes Dev. 2007 Dec 15;21(24):3283-95. doi: 10.1101/gad.1610207. Genes Dev. 2007. PMID: 18079175 Free PMC article.
-
Control of WHITE COLLAR localization by phosphorylation is a critical step in the circadian negative feedback process.EMBO J. 2008 Dec 17;27(24):3246-55. doi: 10.1038/emboj.2008.245. Epub 2008 Nov 20. EMBO J. 2008. PMID: 19020516 Free PMC article.
-
Mechanism of the Neurospora circadian clock, a FREQUENCY-centric view.Biochemistry. 2015 Jan 20;54(2):150-6. doi: 10.1021/bi5005624. Epub 2014 Dec 30. Biochemistry. 2015. PMID: 25302868 Free PMC article. Review.
References
-
- Akten B., Jauch,E., Genova,G.K., Kim,E.Y., Edery,I., Raabe,T. and Jackson,F.R. (2003) A role for CK2 in the Drosophila circadian oscillator. Nat. Neurosci., 6, 251–257. - PubMed
-
- Aronson B., Johnson,K., Loros,J.J. and Dunlap,J.C. (1994a) Negative feedback defining a circadian clock: autoregulation in the clock gene frequency. Science, 263, 1578–1584. - PubMed
-
- Cambareri E.B., Jensen,B.C., Schabtach,E. and Selker,E.U. (1989) Repeat-induced G–C to A–T mutations in Neurospora. Science, 244, 1571–1575. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
