

Progression through key stages of haemopoiesis is dependent on distinct threshold levels of c-Myb
- PMID: 12941699
- PMCID: PMC202376
- DOI: 10.1093/emboj/cdg434
Progression through key stages of haemopoiesis is dependent on distinct threshold levels of c-Myb
Abstract
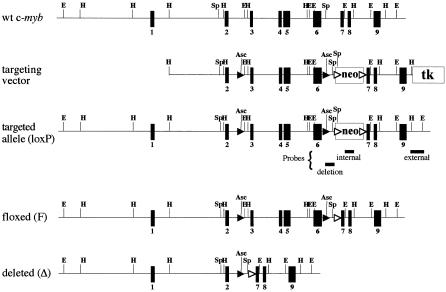
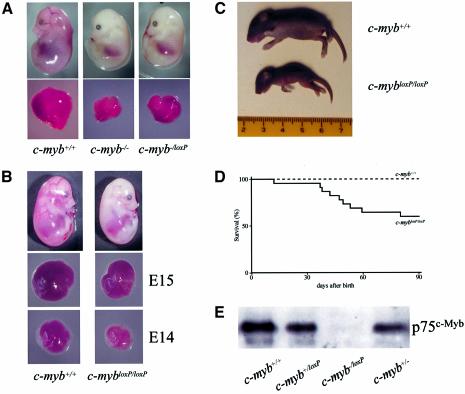
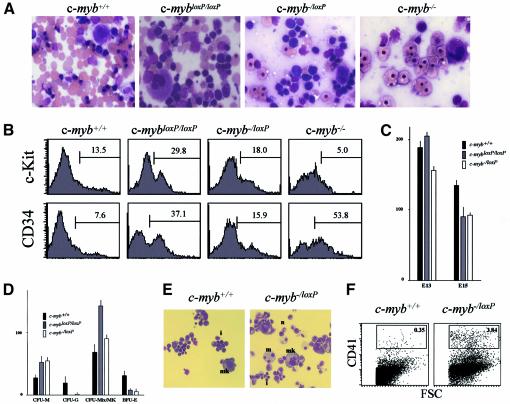
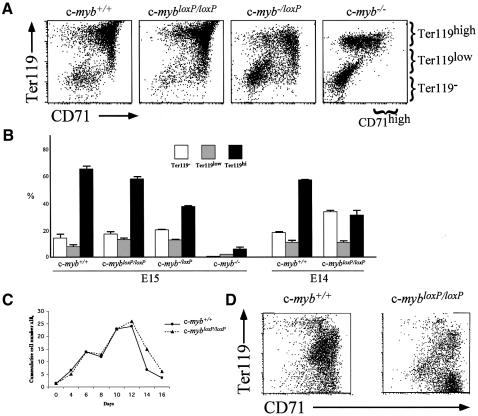
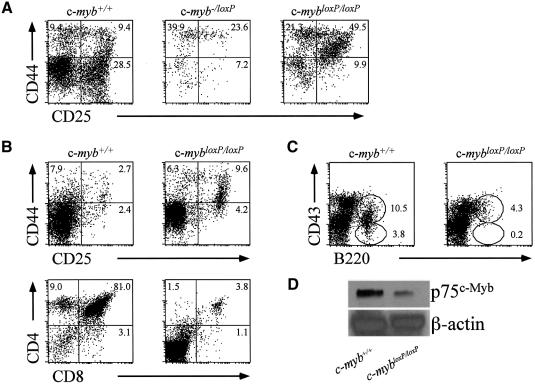
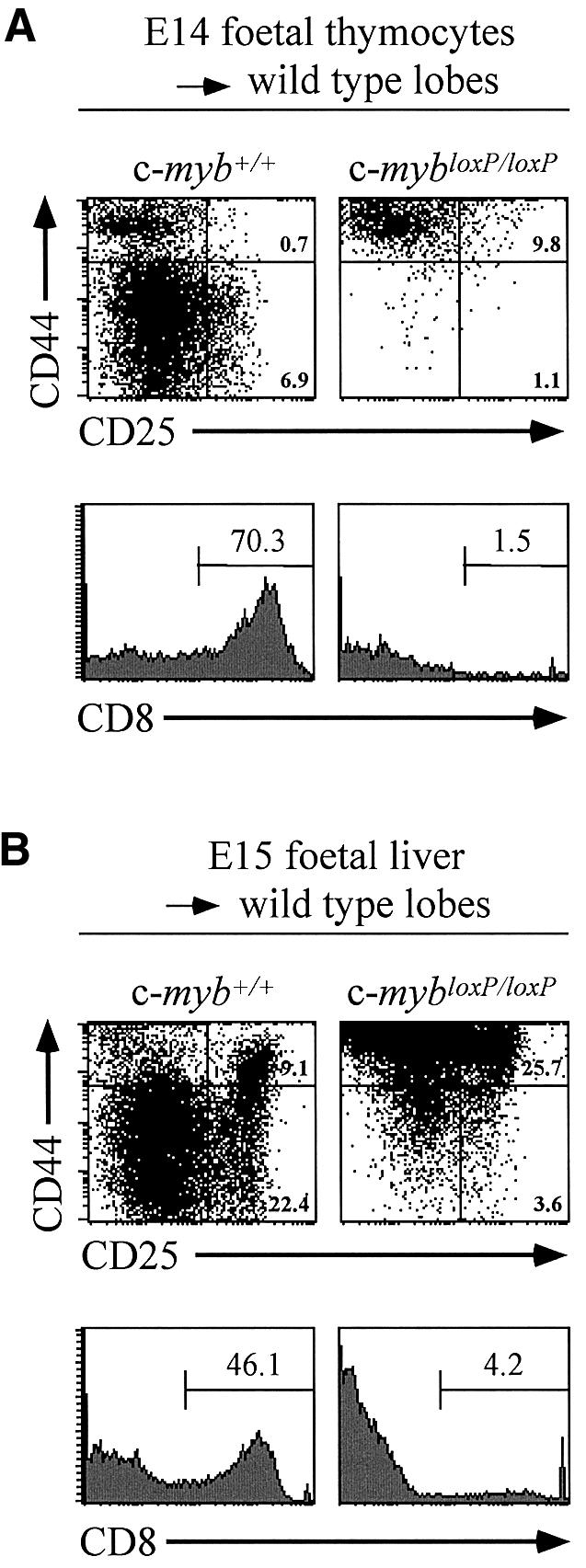
The c-Myb transcription factor is expressed in immature haemopoietic cells and at key stages during differentiation. Loss of the c-myb gene results in embryonic lethality because mature blood cells fail to develop, although commitment to definitive haemopoiesis occurs. We have generated a knockdown allele of c-myb, expressing low levels of the protein, which has enabled us to investigate further the involvement of c-Myb in haemopoiesis. Low levels of c-Myb are sufficient to allow progenitor expansion but, importantly, we show that progression of progenitors towards terminal differentiation is significantly altered. Suboptimal levels of c-Myb favour differentiation of macrophage and megakaryocytes, while higher levels seem to be particularly important in the control of erythropoiesis and lymphopoiesis. We provide evidence that the transition from the CFU-E to erythroblasts is critically dependent on c-Myb levels. During thymopoiesis, c-Myb appears to regulate immature cell numbers and differentiation prior to expression of CD4 and CD8. Overall, our results point to a complex involvement of c-Myb in the regulation of proliferation and commitment within the haemopoietic hierarchy.
Figures
References
-
- Clarke D., Vegiopoulos,A., Crawford,A., Mucenski,M., Bonifer,C. and Frampton,J. (2000) In vitro differentiation of c-myb–/– ES cells reveals that the colony forming capacity of unilineage macrophage precursors and myeloid progenitor commitment are c-Myb independent. Oncogene, 19, 3343–3351. - PubMed
-
- DeKoter R.P. and Singh,H. (2000) Regulation of B lymphocyte and macrophage development by graded expression of PU.1. Science, 288, 1439–1441. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
