

Insulation of the chicken beta-globin chromosomal domain from a chromatin-condensing protein, MENT
- PMID: 12944473
- PMCID: PMC193700
- DOI: 10.1128/MCB.23.18.6455-6468.2003
Insulation of the chicken beta-globin chromosomal domain from a chromatin-condensing protein, MENT
Abstract
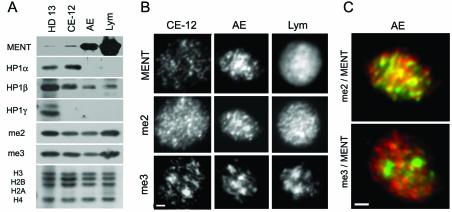
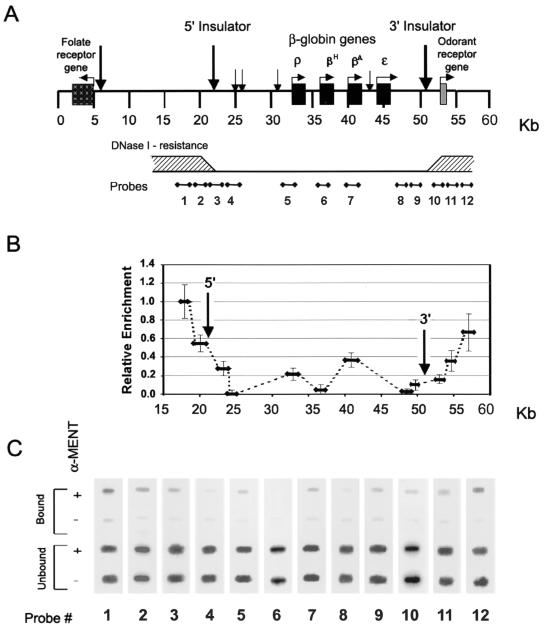
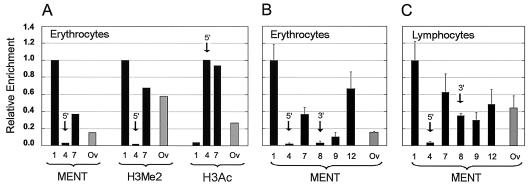
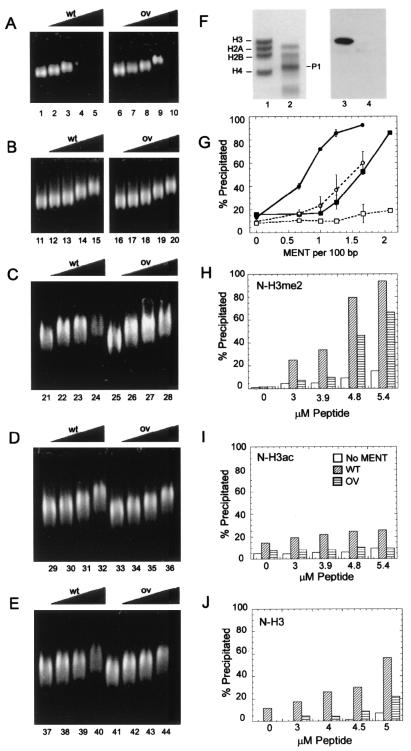
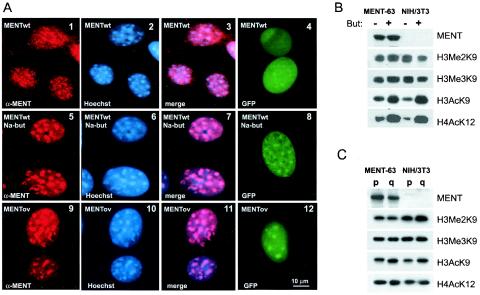
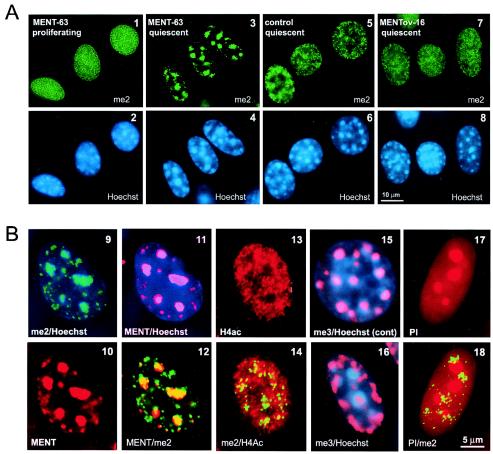
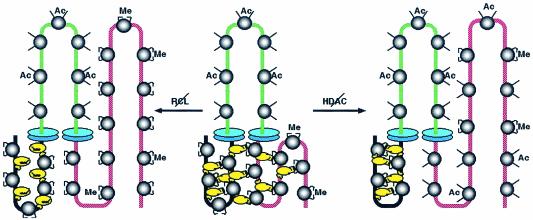
Active genes are insulated from developmentally regulated chromatin condensation in terminally differentiated cells. We mapped the topography of a terminal stage-specific chromatin-condensing protein, MENT, across the active chicken beta-globin domain. We observed two sharp transitions of MENT concentration coinciding with the beta-globin boundary elements. The MENT distribution profile was opposite to that of acetylated core histones but correlated with that of histone H3 dimethylated at lysine 9 (H3me2K9). Ectopic MENT expression in NIH 3T3 cells caused a large-scale and specific remodeling of chromatin marked by H3me2K9. MENT colocalized with H3me2K9 both in chicken erythrocytes and NIH 3T3 cells. Mutational analysis of MENT and experiments with deacetylase inhibitors revealed the essential role of the reaction center loop domain and an inhibitory affect of histone hyperacetylation on the MENT-induced chromatin remodeling in vivo. In vitro, the elimination of the histone H3 N-terminal peptide containing lysine 9 by trypsin blocked chromatin self-association by MENT, while reconstitution with dimethylated but not acetylated N-terminal domain of histone H3 specifically restored chromatin self-association by MENT. We suggest that histone H3 modification at lysine 9 directly regulates chromatin condensation by recruiting MENT to chromatin in a fashion that is spatially constrained from active genes by gene boundary elements and histone hyperacetylation.
Figures
Similar articles
-
Correlation between histone lysine methylation and developmental changes at the chicken beta-globin locus.Science. 2001 Sep 28;293(5539):2453-5. doi: 10.1126/science.1064413. Epub 2001 Aug 9. Science. 2001. PMID: 11498546
-
Stage-specific expression and localization of MENT, a nuclear protein associated with chromatin condensation in terminally differentiating avian erythroid cells.Exp Cell Res. 1993 Jun;206(2):335-43. doi: 10.1006/excr.1993.1154. Exp Cell Res. 1993. PMID: 8500553
-
Chromatin compaction in terminally differentiated avian blood cells: the role of linker histone H5 and non-histone protein MENT.Chromosome Res. 2011 Jul;19(5):579-90. doi: 10.1007/s10577-011-9218-3. Epub 2011 Jun 9. Chromosome Res. 2011. PMID: 21656257 Free PMC article. Review.
-
Histone deacetylase-dependent establishment and maintenance of broad low-level histone acetylation within a tissue-specific chromatin domain.Biochemistry. 2002 Dec 24;41(51):15152-60. doi: 10.1021/bi026786q. Biochemistry. 2002. PMID: 12484752
-
Chromatin domains and regulation of gene expression: familiar and enigmatic clusters of chicken globin genes.Crit Rev Eukaryot Gene Expr. 2001;11(1-3):227-42. Crit Rev Eukaryot Gene Expr. 2001. PMID: 11693962 Review.
Cited by
-
Cathepsin L stabilizes the histone modification landscape on the Y chromosome and pericentromeric heterochromatin.Mol Cell Biol. 2006 Jun;26(11):4172-84. doi: 10.1128/MCB.00135-06. Mol Cell Biol. 2006. PMID: 16705169 Free PMC article.
-
BAZ1B the Protean Protein.Genes (Basel). 2021 Sep 28;12(10):1541. doi: 10.3390/genes12101541. Genes (Basel). 2021. PMID: 34680936 Free PMC article. Review.
-
Developmentally regulated linker histone H1c promotes heterochromatin condensation and mediates structural integrity of rod photoreceptors in mouse retina.J Biol Chem. 2013 Jun 14;288(24):17895-907. doi: 10.1074/jbc.M113.452144. Epub 2013 May 3. J Biol Chem. 2013. PMID: 23645681 Free PMC article.
-
Insights into HP1a-Chromatin Interactions.Cells. 2020 Aug 9;9(8):1866. doi: 10.3390/cells9081866. Cells. 2020. PMID: 32784937 Free PMC article. Review.
-
Heterochromatin protein 1alpha: a hallmark of cell proliferation relevant to clinical oncology.EMBO Mol Med. 2009 Jun;1(3):178-91. doi: 10.1002/emmm.200900022. EMBO Mol Med. 2009. PMID: 20049717 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources