

The checkpoint protein Rad24 of Saccharomyces cerevisiae is involved in processing double-strand break ends and in recombination partner choice
- PMID: 12944484
- PMCID: PMC193701
- DOI: 10.1128/MCB.23.18.6585-6596.2003
The checkpoint protein Rad24 of Saccharomyces cerevisiae is involved in processing double-strand break ends and in recombination partner choice
Abstract
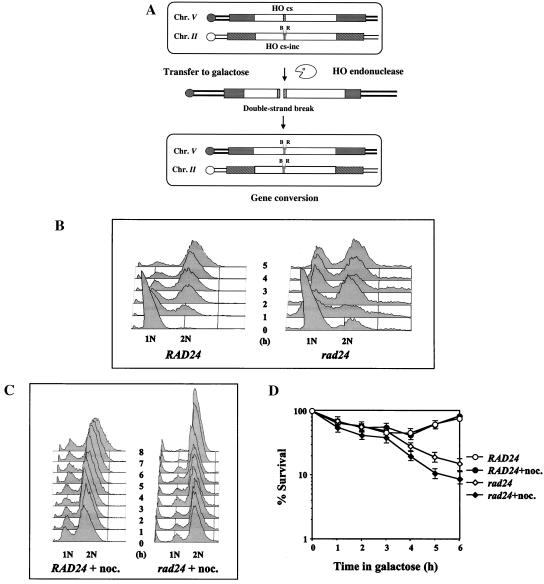
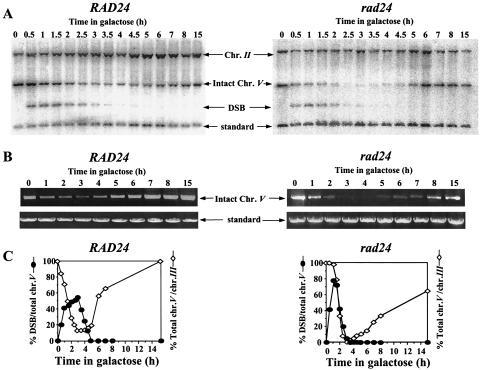
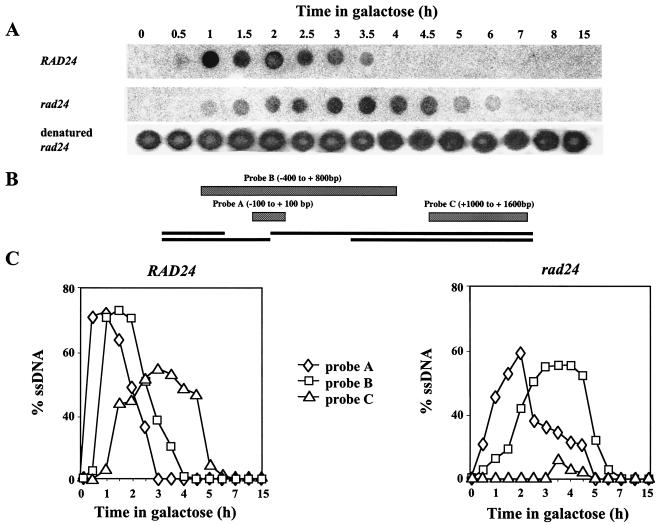
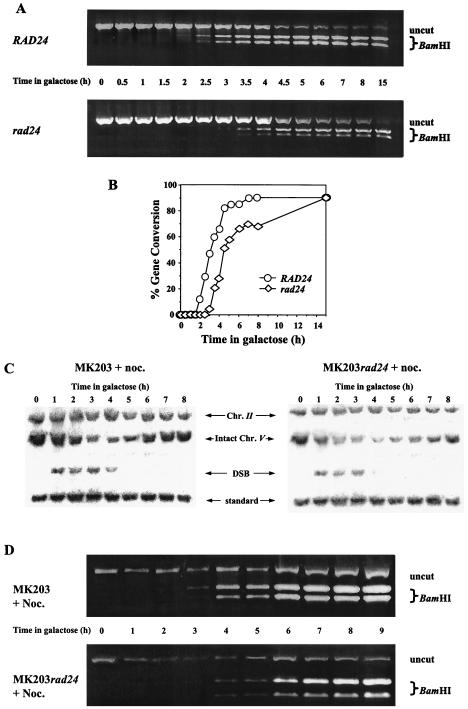
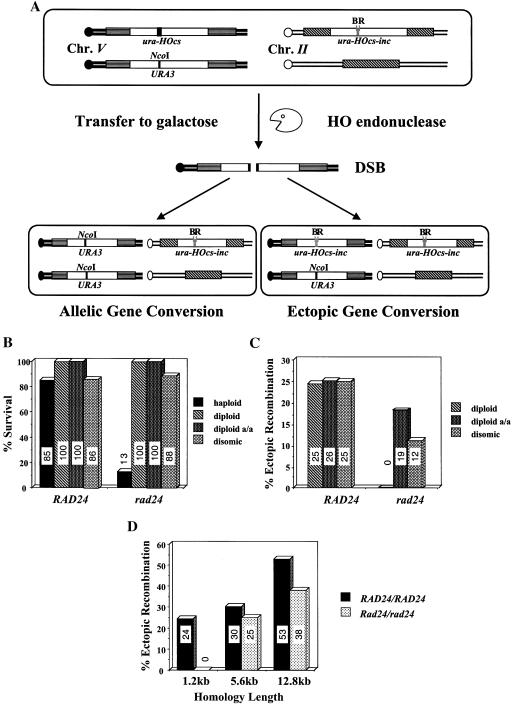
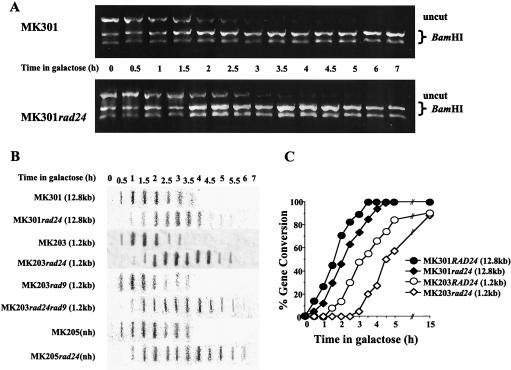
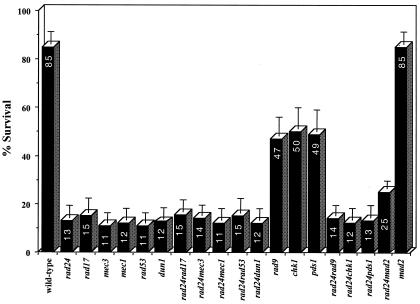
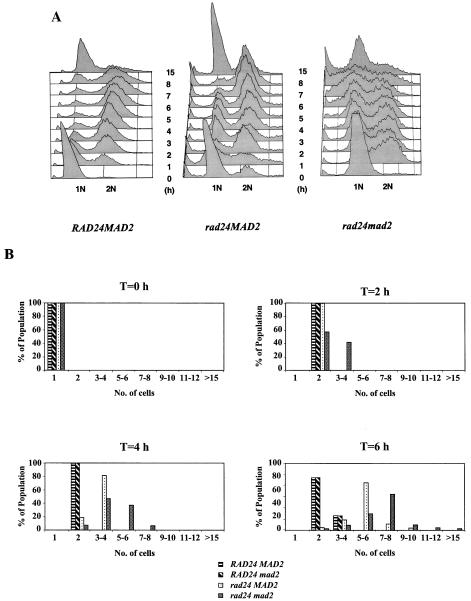
Upon chromosomal damage, cells activate a checkpoint response that includes cell cycle arrest and a stimulation of DNA repair. The checkpoint protein Rad24 is key to the survival of a single, repairable double-strand break (DSB). However, the low survival of rad24 cells is not due to their inability to arrest cell cycle progression. In rad24 mutants, processing of the broken ends is delayed and protracted, resulting in extended kinetics of DSB repair and in cell death. The limited resection of rad24 mutants also affects recombination partner choice by a mechanism dependent on the length of the interacting homologous donor sequences. Unexpectedly, rad24 cells with a DSB eventually accumulate and die at the G(2)/M phase of the cell cycle. This arrest depends on the spindle checkpoint protein Mad2.
Figures
References
-
- Carr, A. M. 1997. Control of cell cycle arrest by the Mec1sc/Rad3sp DNA structure checkpoint pathway. Curr. Opin. Genet. Deversion 7:93-98. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases