

Active self-splicing group I introns in 23S rRNA genes of hyperthermophilic bacteria, derived from introns in eukaryotic organelles
- PMID: 12947037
- PMCID: PMC196884
- DOI: 10.1073/pnas.1434268100
Active self-splicing group I introns in 23S rRNA genes of hyperthermophilic bacteria, derived from introns in eukaryotic organelles
Erratum in
- Proc Natl Acad Sci U S A. 2003 Nov 25;100(24):14511
Abstract
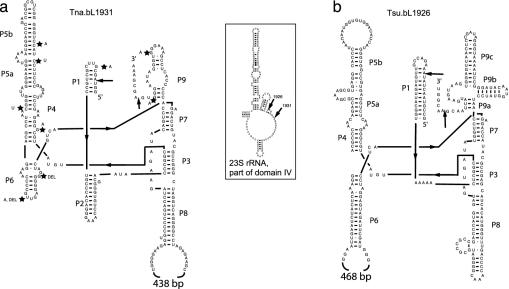
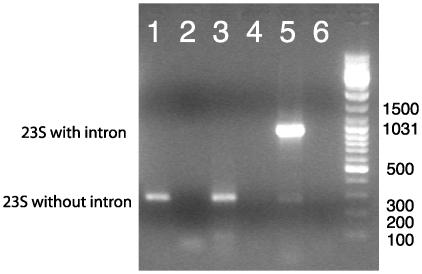
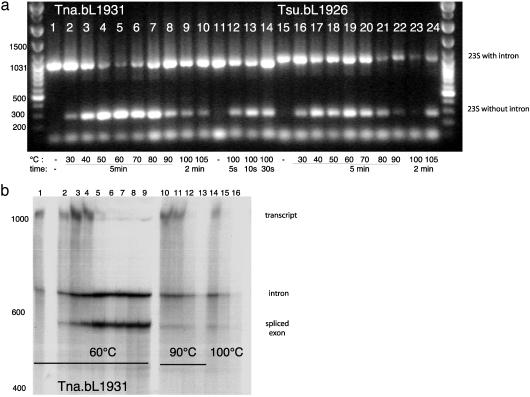
Group I introns are common in the 23 rRNA genes of mitochondria and chloroplasts. Often, they encode "homing endonucleases," which target highly conserved gene sequences and drive interorganellar intron mobility, even across species and genus lines. Most bacterial 23S rRNA genes show these same endonuclease-sensitive target sequences. However, only two bacterial 23S rRNA genes are known to contain group I introns: that of Simkania negevensis [Everett, K. D., Kahane, S., Bush, R. M. & Friedman, M. G. (1999) J. Bacteriol. 181, 4734-4740], where the intron is not spliced and probably limits growth, and that of Coxiella burnetii [Seshadri, R., et al. (2003) Proc. Natl. Acad. Sci. USA 100, 5455-5460], where no direct evidence of splicing exists. Both bacteria are intracellular parasites and might have acquired introns from eukaryotic hosts. Here we provide direct evidence for splicing, and evolutionary evidence for mobility, of group I introns in the 23S rRNA genes of several free-living hyperthermophilic bacteria of the genus Thermotoga. These bacteria do not live closely with eukaryotes, but phylogenetic analyses suggest that their introns were also acquired from eukaryotic (probably algal) organelles. In vivo, their introns must be spliced at temperatures approaching 90 degrees C, making them the most thermostable natural ribozymes so far described. We demonstrate that at least some of these introns can also self-splice in vitro.
Figures


References
-
- Cech, T. R. (1990) Annu. Rev. Biochem. 59, 543–568. - PubMed
-
- Cech, T. R. (2001) Biochem. Soc. Trans. 30, 1162–1166. - PubMed
-
- Lambowitz, A. M. & Belfort, M. (1993) Annu. Rev. Biochem. 62, 587–622. - PubMed
-
- Jackson, S., Cannone, J., Lee, J., Gutell, R. & Woodson, S. (2002) J. Mol. Biol. 323, 35–52. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
