

Cdyl: a new transcriptional co-repressor
- PMID: 12947414
- PMCID: PMC1326355
- DOI: 10.1038/sj.embor.embor917
Cdyl: a new transcriptional co-repressor
Abstract
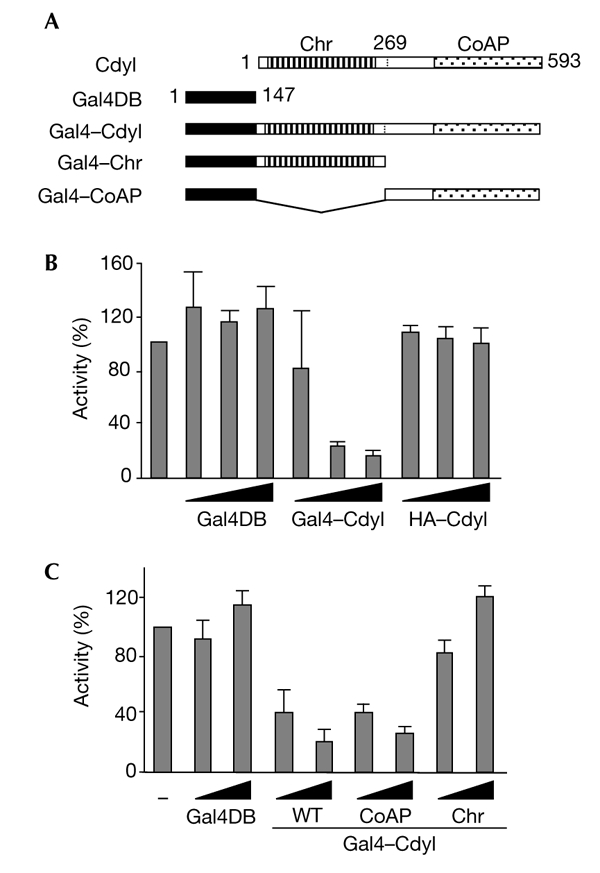
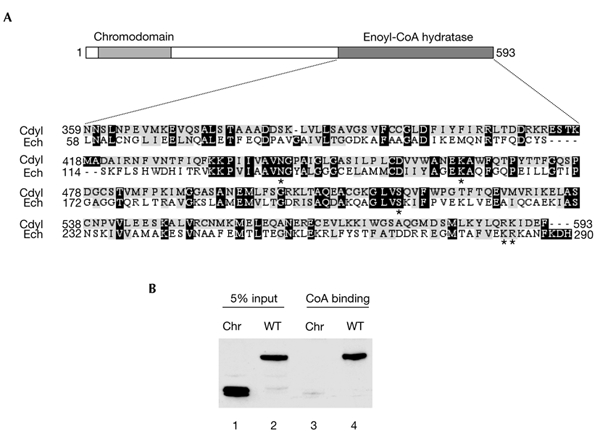
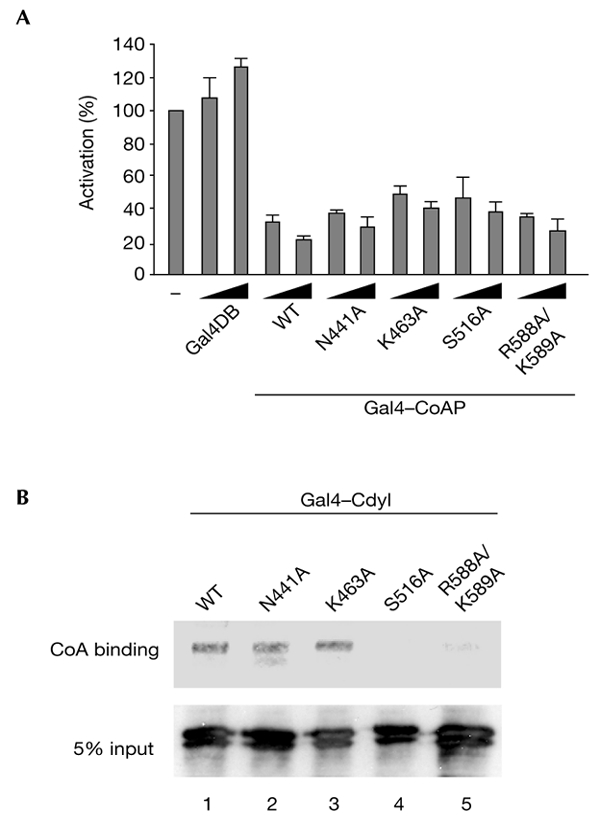
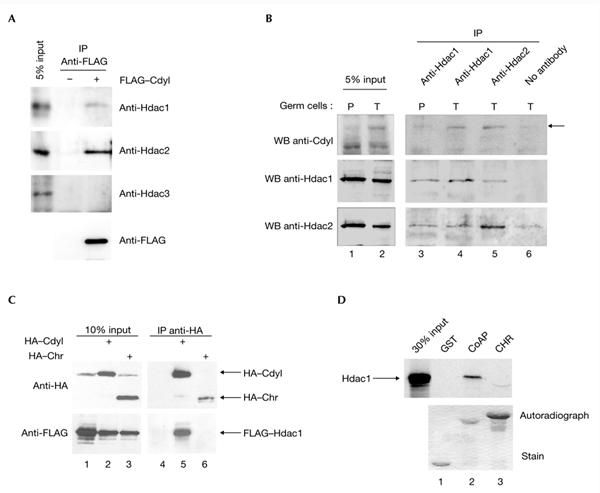
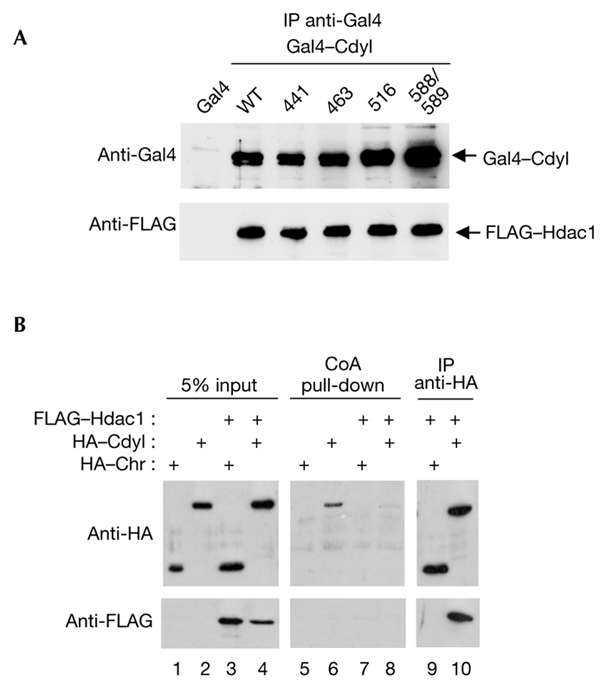
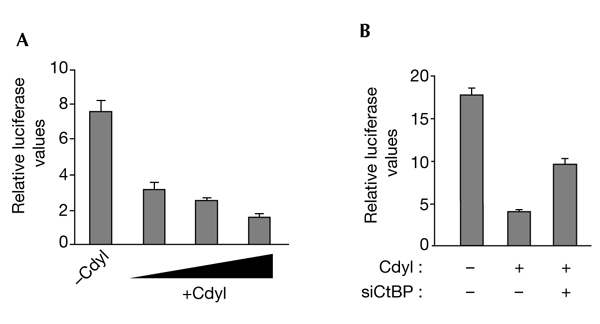
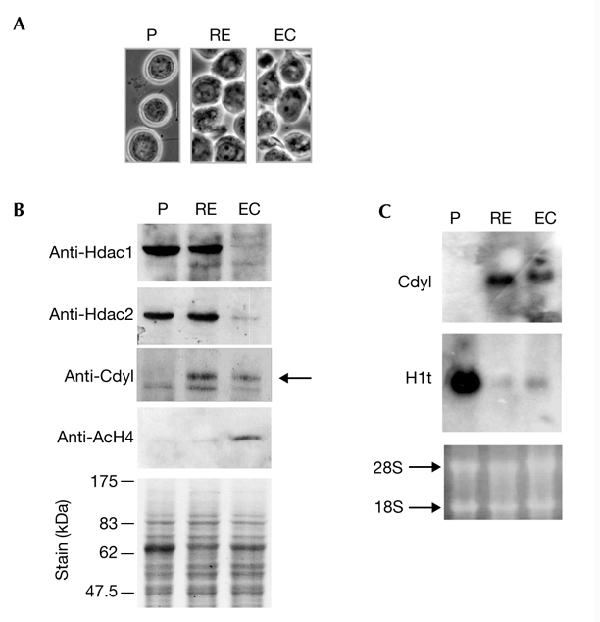
Cdyl (chromodomain-Y-like) is a chromodomain-containing protein that is predominantly expressed during mouse spermiogenesis. In its carboxy-terminal portion, there is a domain with homology to the coenzyme A (CoA) pocket of the enoyl-CoA hydratase/isomerase, which is shown here to be able to bind CoA and histone deacetylases (HDACs). It also efficiently represses transcription. Moreover, the binding of Hdac1 represses the ability of Cdyl to bind CoA, and a Cdyl-CoA interaction only occurs in the absence of HDACs. These data suggest that Cdyl is primarily a transcriptional co-repressor. However, the degradation of cellular Hdac1 and Hdac2, as observed here in the elongating spermatids, may provide an HDAC-free environment in which Cdyl could bind CoA and participate in the global chromatin remodelling that occurs in these cells.
Figures
References
-
- Bellve A.R. ( 1993) Purification, culture, and fractionation of spermatogenic cells. Methods Enzymol., 225, 84–113. - PubMed
-
- Comijn J., Berx G., Vermassen P., Verschueren K., van Grunsven L.A., Bruyneel E., Mareel M., Huylebroek D. & van Roy F. ( 2001) The two handed E-box binding zinc finger protein SIP1 downregulates E-cadherin and induces invasion. Mol. Cell, 7, 1267–1278. - PubMed
-
- Lahn B.T. & Page D.C. ( 1999) Retroposition of autosomal mRNA yielded testis-specific gene family on human Y chromosome. Nature Genet., 21, 429–433. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
