

Effect of translational signals on mRNA decay in Bacillus subtilis
- PMID: 12949089
- PMCID: PMC193772
- DOI: 10.1128/JB.185.18.5372-5379.2003
Effect of translational signals on mRNA decay in Bacillus subtilis
Abstract
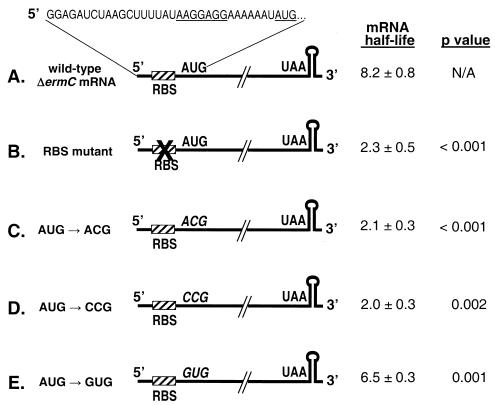
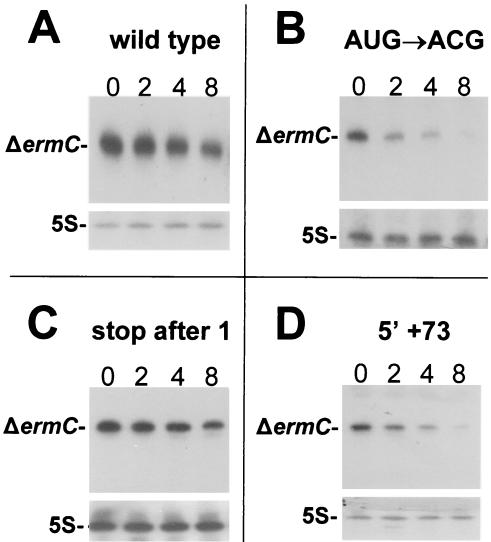
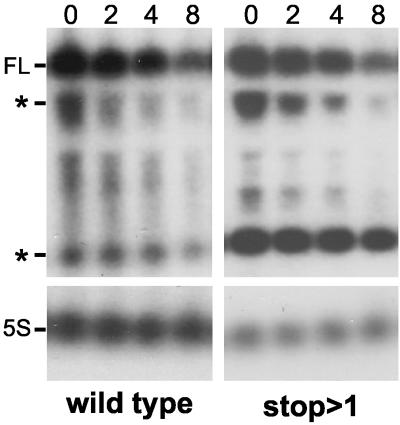
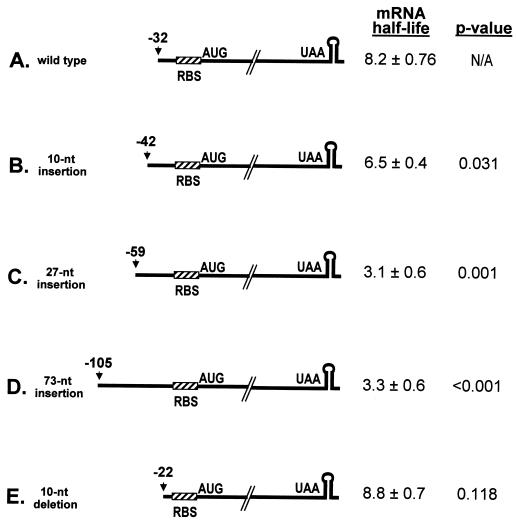
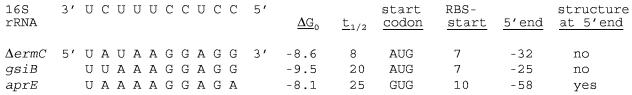
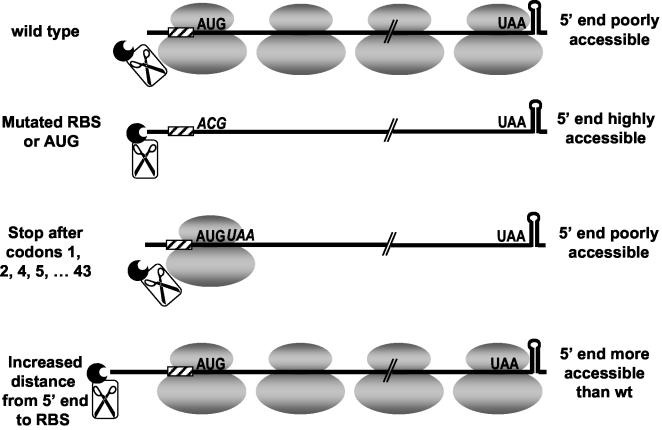
A 254-nucleotide model mRNA, designated deltaermC mRNA, was used to study the effects of translational signals and ribosome transit on mRNA decay in Bacillus subtilis. DeltaermC mRNA features a strong ribosome-binding site (RBS) and a 62-amino-acid-encoding open reading frame, followed by a transcription terminator structure. Inactivation of the RBS or the start codon resulted in a fourfold decrease in the mRNA half-life, demonstrating the importance of ternary complex formation for mRNA stability. Data for the decay of deltaermC mRNAs with stop codons at positions increasingly proximal to the translational start site showed that actual translation--even the formation of the first peptide bond--was not important for stability. The half-life of an untranslated 3.2-kb deltaermC-lacZ fusion RNA was similar to that of a translated deltaermC-lacZ mRNA, indicating that the translation of even a longer RNA was not required for wild-type stability. The data are consistent with a model in which ribosome binding and the formation of the ternary complex interfere with a 5'-end-dependent activity, possibly a 5'-binding endonuclease, which is required for the initiation of mRNA decay. This model is supported by the finding that increasing the distance from the 5' end to the start codon resulted in a 2.5-fold decrease in the mRNA half-life. These results underscore the importance of the 5' end to mRNA stability in B. subtilis.
Figures

Similar articles
-
Erythromycin-induced ribosome stalling and RNase J1-mediated mRNA processing in Bacillus subtilis.Mol Microbiol. 2008 Sep;69(6):1439-49. doi: 10.1111/j.1365-2958.2008.06370.x. Epub 2008 Jul 18. Mol Microbiol. 2008. PMID: 18647167 Free PMC article.
-
Bacillus subtilis RNase J1 endonuclease and 5' exonuclease activities in the turnover of DeltaermC mRNA.RNA. 2009 Dec;15(12):2331-9. doi: 10.1261/rna.1749109. Epub 2009 Oct 22. RNA. 2009. PMID: 19850915 Free PMC article.
-
A 5' stem-loop and ribosome binding but not translation are important for the stability of Bacillus subtilis aprE leader mRNA.Microbiology (Reading). 2002 Jun;148(Pt 6):1795-1803. doi: 10.1099/00221287-148-6-1795. Microbiology (Reading). 2002. PMID: 12055299
-
Ribosome dynamics and mRNA turnover, a complex relationship under constant cellular scrutiny.Wiley Interdiscip Rev RNA. 2021 Nov;12(6):e1658. doi: 10.1002/wrna.1658. Epub 2021 May 5. Wiley Interdiscip Rev RNA. 2021. PMID: 33949788 Free PMC article. Review.
-
Messenger RNA decay and maturation in Bacillus subtilis.Prog Mol Biol Transl Sci. 2009;85:231-73. doi: 10.1016/S0079-6603(08)00806-4. Prog Mol Biol Transl Sci. 2009. PMID: 19215774 Review.
Cited by
-
The Effect of an Alternate Start Codon on Heterologous Expression of a PhoA Fusion Protein in Mycoplasma gallisepticum.PLoS One. 2015 May 26;10(5):e0127911. doi: 10.1371/journal.pone.0127911. eCollection 2015. PLoS One. 2015. PMID: 26010086 Free PMC article.
-
Ribonucleases J1 and J2: two novel endoribonucleases in B.subtilis with functional homology to E.coli RNase E.Nucleic Acids Res. 2005 Apr 14;33(7):2141-52. doi: 10.1093/nar/gki505. Print 2005. Nucleic Acids Res. 2005. PMID: 15831787 Free PMC article.
-
Composability of regulatory sequences controlling transcription and translation in Escherichia coli.Proc Natl Acad Sci U S A. 2013 Aug 20;110(34):14024-9. doi: 10.1073/pnas.1301301110. Epub 2013 Aug 7. Proc Natl Acad Sci U S A. 2013. PMID: 23924614 Free PMC article.
-
Decay of a model mRNA in Bacillus subtilis by a combination of RNase J1 5' exonuclease and RNase Y endonuclease activities.J Bacteriol. 2011 Nov;193(22):6384-6. doi: 10.1128/JB.05939-11. Epub 2011 Sep 9. J Bacteriol. 2011. PMID: 21908660 Free PMC article.
-
Expression of multiple Bacillus subtilis genes is controlled by decay of slrA mRNA from Rho-dependent 3' ends.Nucleic Acids Res. 2016 Apr 20;44(7):3364-72. doi: 10.1093/nar/gkw069. Epub 2016 Feb 8. Nucleic Acids Res. 2016. PMID: 26857544 Free PMC article.
References
-
- Bechhofer, D. H. 1993. 5′ mRNA stabilizers, p. 31-52. In J. G. Belasco and G. Brawerman (ed.), Control of mRNA stability. Academic Press, Inc., San Diego, Calif.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
