

Moritella cold-active dihydrofolate reductase: are there natural limits to optimization of catalytic efficiency at low temperature?
- PMID: 12949104
- PMCID: PMC193739
- DOI: 10.1128/JB.185.18.5519-5526.2003
Moritella cold-active dihydrofolate reductase: are there natural limits to optimization of catalytic efficiency at low temperature?
Abstract
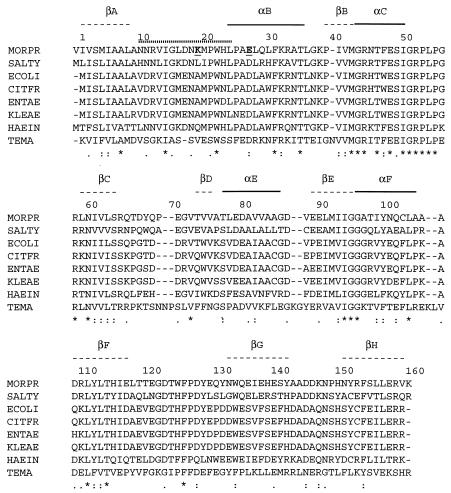
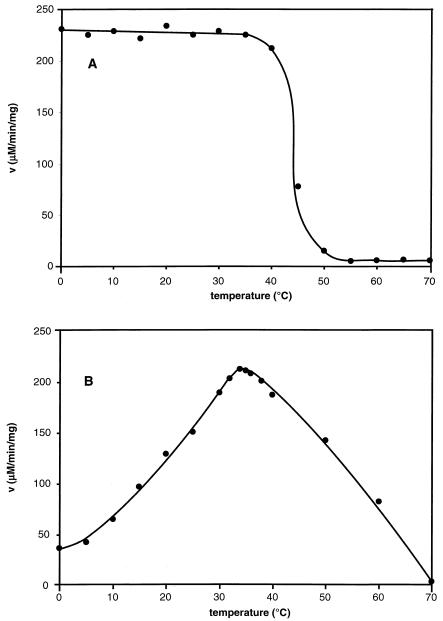
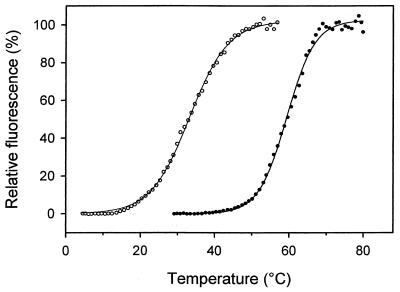
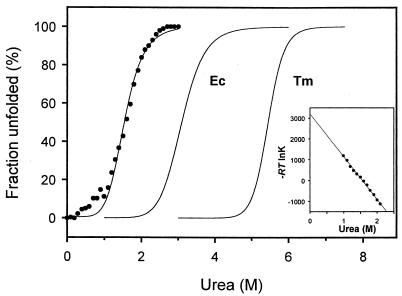
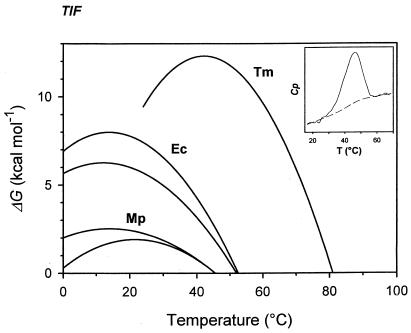
Adapting metabolic enzymes of microorganisms to low temperature environments may require a difficult compromise between velocity and affinity. We have investigated catalytic efficiency in a key metabolic enzyme (dihydrofolate reductase) of Moritella profunda sp. nov., a strictly psychrophilic bacterium with a maximal growth rate at 2 degrees C or less. The enzyme is monomeric (Mr=18,291), 55% identical to its Escherichia coli counterpart, and displays Tm and denaturation enthalpy changes much lower than E. coli and Thermotoga maritima homologues. Its stability curve indicates a maximum stability above the temperature range of the organism, and predicts cold denaturation below 0 degrees C. At mesophilic temperatures the apparent Km value for dihydrofolate is 50- to 80-fold higher than for E. coli, Lactobacillus casei, and T. maritima dihydrofolate reductases, whereas the apparent Km value for NADPH, though higher, remains in the same order of magnitude. At 5 degrees C these values are not significantly modified. The enzyme is also much less sensitive than its E. coli counterpart to the inhibitors methotrexate and trimethoprim. The catalytic efficiency (kcat/Km) with respect to dihydrofolate is thus much lower than in the other three bacteria. The higher affinity for NADPH could have been maintained by selection since NADPH assists the release of the product tetrahydrofolate. Dihydrofolate reductase adaptation to low temperature thus appears to have entailed a pronounced trade-off between affinity and catalytic velocity. The kinetic features of this psychrophilic protein suggest that enzyme adaptation to low temperature may be constrained by natural limits to optimization of catalytic efficiency.
Figures
Similar articles
-
Reduced susceptibility of Moritella profunda dihydrofolate reductase to trimethoprim is not due to glutamate 28.Protein J. 2011 Dec;30(8):546-8. doi: 10.1007/s10930-011-9361-x. Protein J. 2011. PMID: 21968646
-
Site-directed mutagenesis of mouse dihydrofolate reductase. Mutants with increased resistance to methotrexate and trimethoprim.J Biol Chem. 1988 Sep 5;263(25):12500-8. J Biol Chem. 1988. PMID: 3045118
-
Metabolic enzymes from psychrophilic bacteria: challenge of adaptation to low temperatures in ornithine carbamoyltransferase from Moritella abyssi.J Bacteriol. 2003 Apr;185(7):2161-8. doi: 10.1128/JB.185.7.2161-2168.2003. J Bacteriol. 2003. PMID: 12644485 Free PMC article.
-
Environmental Adaptation of Dihydrofolate Reductase from Deep-Sea Bacteria.Subcell Biochem. 2015;72:423-42. doi: 10.1007/978-94-017-9918-8_21. Subcell Biochem. 2015. PMID: 26174394 Review.
-
Optimization to low temperature activity in psychrophilic enzymes.Int J Mol Sci. 2012;13(9):11643-11665. doi: 10.3390/ijms130911643. Epub 2012 Sep 17. Int J Mol Sci. 2012. PMID: 23109875 Free PMC article. Review.
Cited by
-
Adaptations for Pressure and Temperature in Dihydrofolate Reductases.Microorganisms. 2021 Aug 11;9(8):1706. doi: 10.3390/microorganisms9081706. Microorganisms. 2021. PMID: 34442785 Free PMC article.
-
Dihydropteridine reductase as an alternative to dihydrofolate reductase for synthesis of tetrahydrofolate in Thermus thermophilus.J Bacteriol. 2004 Jan;186(2):351-5. doi: 10.1128/JB.186.2.351-355.2004. J Bacteriol. 2004. PMID: 14702303 Free PMC article.
-
Continuous selection pressure to improve temperature acclimation of Tisochrysis lutea.PLoS One. 2017 Sep 13;12(9):e0183547. doi: 10.1371/journal.pone.0183547. eCollection 2017. PLoS One. 2017. PMID: 28902878 Free PMC article.
-
A diminished hydrophobic effect inside the GroEL/ES cavity contributes to protein substrate destabilization.Proc Natl Acad Sci U S A. 2022 Nov 29;119(48):e2213170119. doi: 10.1073/pnas.2213170119. Epub 2022 Nov 21. Proc Natl Acad Sci U S A. 2022. PMID: 36409898 Free PMC article.
-
Comparative study on dihydrofolate reductases from Shewanella species living in deep-sea and ambient atmospheric-pressure environments.Extremophiles. 2011 Mar;15(2):165-75. doi: 10.1007/s00792-010-0345-0. Epub 2010 Dec 23. Extremophiles. 2011. PMID: 21181485
References
-
- Baccanari, D. P., D. Stone, and L. Kuyper. 1981. Effect of a single amino acid substitution on Escherichia coli dihydrofolate reductase catalysis and ligand binding. J. Biol. Chem. 256:1738-1747. - PubMed
-
- Bentahir, M., G. Feller, M. Aittaleb, J. Lamotte-Brasseur, T. Himri, J. P. Chessa, and C. Gerday. 2000. Structural, kinetic and calorimetric characterization of the cold-active phosphoglycerate kinase from the antarctic Pseudomonas sp. TACII18. J. Biol. Chem. 275:11147-11153. - PubMed
-
- Bolin, J. T., D. J. Filman, D. A. Matthews, R. C. Hamlin, and J. Kraut. 1982. Crystal structures of Escherichia coli and Lactobacillus casei dihydrofolate reductase refined at 1.7 A resolution. I. General features and binding of methotrexate. J. Biol. Chem. 257:13650-13662. - PubMed
-
- Cavicchioli, R., K. S. Siddiqui, D. Andrews, and R. K. Sowers. 2002. Low-temperature extremophiles and their applications. Curr. Opin. Biotechnol. 13:253-261. - PubMed
-
- Clark, A. C., and C. Frieden. 1999. Native Escherichia coli and murine dihydrofolate reductases contain late-folding non-native structures. J. Mol. Biol. 285:1765-1776. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
