

Physiological studies of Escherichia coli strain MG1655: growth defects and apparent cross-regulation of gene expression
- PMID: 12949114
- PMCID: PMC193769
- DOI: 10.1128/JB.185.18.5611-5626.2003
Physiological studies of Escherichia coli strain MG1655: growth defects and apparent cross-regulation of gene expression
Abstract
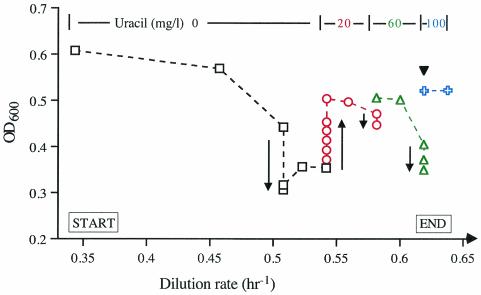
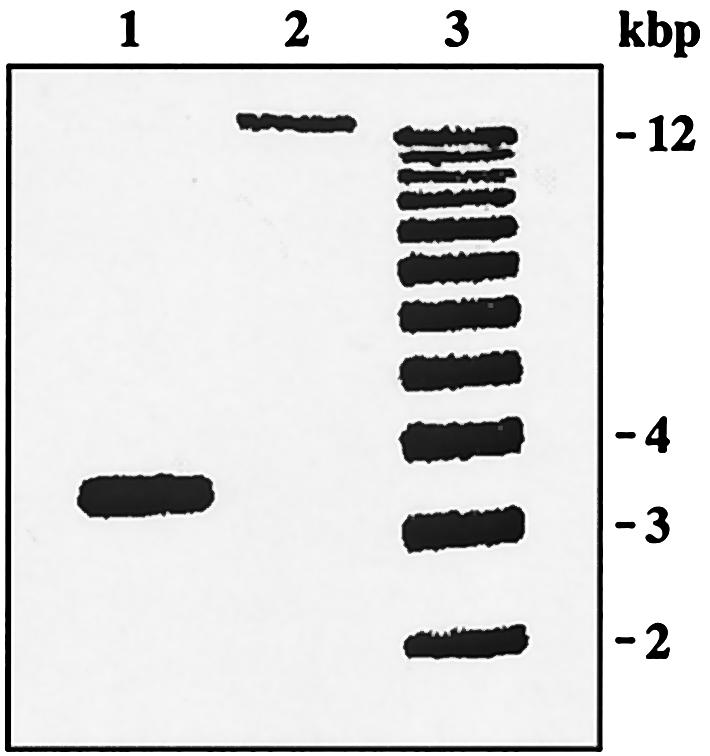
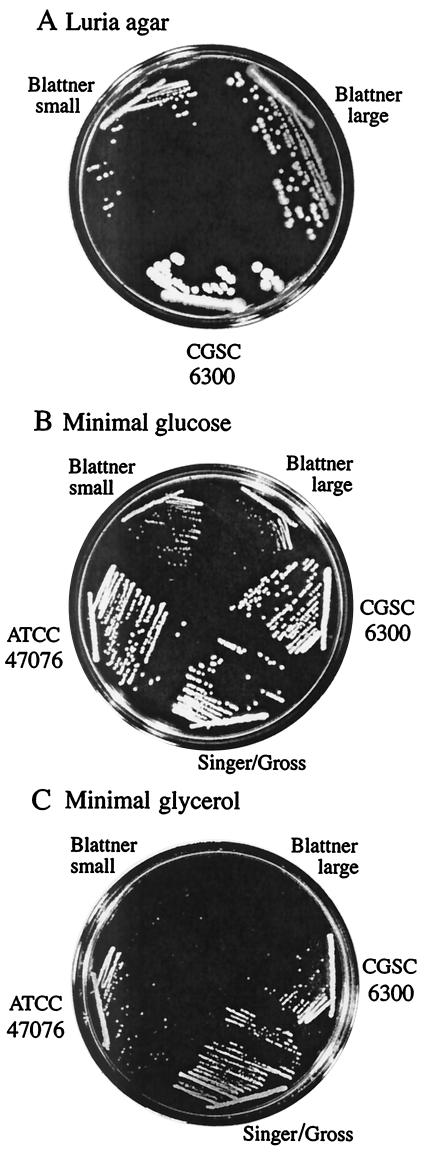
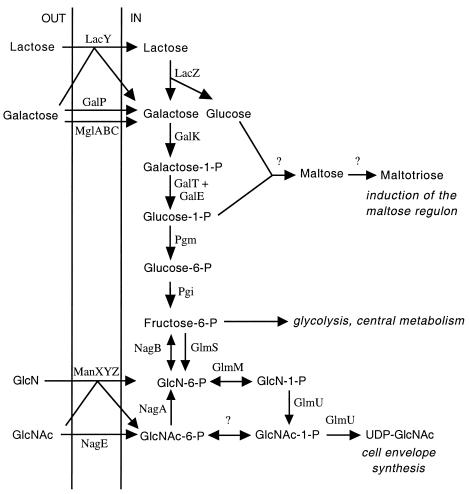
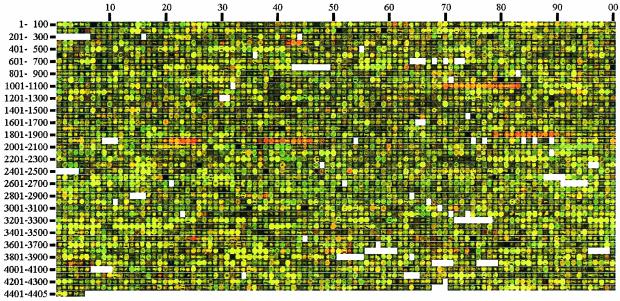
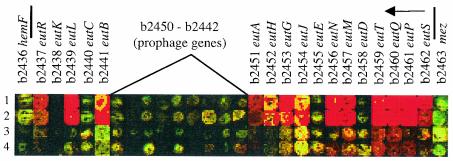
Escherichia coli strain MG1655 was chosen for sequencing because the few mutations it carries (ilvG rfb-50 rph-1) were considered innocuous. However, it has a number of growth defects. Internal pyrimidine starvation due to polarity of the rph-1 allele on pyrE was problematic in continuous culture. Moreover, the isolate of MG1655 obtained from the E. coli Genetic Stock Center also carries a large deletion around the fnr (fumarate-nitrate respiration) regulatory gene. Although studies on DNA microarrays revealed apparent cross-regulation of gene expression between galactose and lactose metabolism in the Stock Center isolate of MG1655, this was due to the occurrence of mutations that increased lacY expression and suppressed slow growth on galactose. The explanation for apparent cross-regulation between galactose and N-acetylglucosamine metabolism was similar. By contrast, cross-regulation between lactose and maltose metabolism appeared to be due to generation of internal maltosaccharides in lactose-grown cells and may be physiologically significant. Lactose is of restricted distribution: it is normally found together with maltosaccharides, which are starch degradation products, in the mammalian intestine. Strains designated MG1655 and obtained from other sources differed from the Stock Center isolate and each other in several respects. We confirmed that use of other E. coli strains with MG1655-based DNA microarrays works well, and hence these arrays can be used to study any strain of interest. The responses to nitrogen limitation of two urinary tract isolates and an intestinal commensal strain isolated recently from humans were remarkably similar to those of MG1655.
Figures

Similar articles
-
[Construction of the new Escherichia coli K-12 wild-type strain with improved growth characteristics for application in metabolic engineering].Genetika. 2010 Mar;46(3):349-55. Genetika. 2010. PMID: 20391779 Russian.
-
Microarray analysis of gene regulation by oxygen, nitrate, nitrite, FNR, NarL and NarP during anaerobic growth of Escherichia coli: new insights into microbial physiology.Biochem Soc Trans. 2006 Feb;34(Pt 1):104-7. doi: 10.1042/BST0340104. Biochem Soc Trans. 2006. PMID: 16417494
-
A reassessment of the FNR regulon and transcriptomic analysis of the effects of nitrate, nitrite, NarXL, and NarQP as Escherichia coli K12 adapts from aerobic to anaerobic growth.J Biol Chem. 2006 Feb 24;281(8):4802-15. doi: 10.1074/jbc.M512312200. Epub 2005 Dec 22. J Biol Chem. 2006. PMID: 16377617
-
The Escherichia coli K-12 "wild types" W3110 and MG1655 have an rph frameshift mutation that leads to pyrimidine starvation due to low pyrE expression levels.J Bacteriol. 1993 Jun;175(11):3401-7. doi: 10.1128/jb.175.11.3401-3407.1993. J Bacteriol. 1993. PMID: 8501045 Free PMC article.
-
The galactose regulon of Escherichia coli.Mol Microbiol. 1993 Oct;10(2):245-51. doi: 10.1111/j.1365-2958.1993.tb01950.x. Mol Microbiol. 1993. PMID: 7934815 Review.
Cited by
-
Complete Genome Sequence of Escherichia coli NCM3722.Genome Announc. 2015 Aug 6;3(4):e00879-15. doi: 10.1128/genomeA.00879-15. Genome Announc. 2015. PMID: 26251500 Free PMC article.
-
A universal trade-off between growth and lag in fluctuating environments.Nature. 2020 Aug;584(7821):470-474. doi: 10.1038/s41586-020-2505-4. Epub 2020 Jul 15. Nature. 2020. PMID: 32669712 Free PMC article.
-
Growth of wildtype and mutant E. coli strains in minimal media for optimal production of nucleic acids for preparing labeled nucleotides.Appl Microbiol Biotechnol. 2010 Oct;88(3):771-9. doi: 10.1007/s00253-010-2813-y. Epub 2010 Aug 21. Appl Microbiol Biotechnol. 2010. PMID: 20730533 Free PMC article.
-
Heterogeneous absorption of antimicrobial peptide LL37 in Escherichia coli cells enhances population survivability.Elife. 2018 Dec 18;7:e38174. doi: 10.7554/eLife.38174. Elife. 2018. PMID: 30560784 Free PMC article.
-
Adaptive prediction of environmental changes by microorganisms.Nature. 2009 Jul 9;460(7252):220-4. doi: 10.1038/nature08112. Epub 2009 Jun 17. Nature. 2009. PMID: 19536156
References
-
- Bachman, B. J. 1996. Derivations and genotypes of some mutant derivatives of Escherichia coli K-12, p. 2460-2488. In F. C. Neidhardt et al. (ed.), Escherichia coli and Salmonella: cellular and molecular biology, 2nd ed., vol. 2. ASM Press, Washington, D.C.
-
- Blattner, F. R., G. Plunkett, 3rd, C. A. Bloch, N. T. Perna, V. Burland, M. Riley, J. Collado-Vides, J. D. Glasner, C. K. Rode, G. F. Mayhew, J. Gregor, N. W. Davis, H. A. Kirkpatrick, M. A. Goeden, D. J. Rose, B. Mau, and Y. Shao. 1997. The complete genome sequence of Escherichia coli K-12. Science 277:1453-1474. - PubMed
-
- Blomfield, I. C., V. Vaughn, R. F. Rest, and B. I. Eisenstein. 1991. Allelic exchange in Escherichia coli with the Bacillus subtilis sacB gene and a temperature-sensitive pSC101 replicon. Mol. Microbiol. 5:1447-1458. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
