

Molecular biology of feather morphogenesis: a testable model for evo-devo research
- PMID: 12949772
- PMCID: PMC4382008
- DOI: 10.1002/jez.b.29
Molecular biology of feather morphogenesis: a testable model for evo-devo research
Abstract
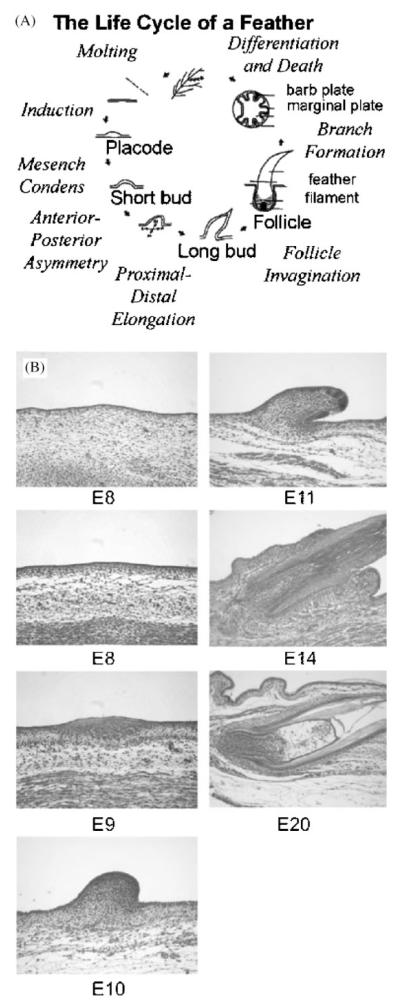
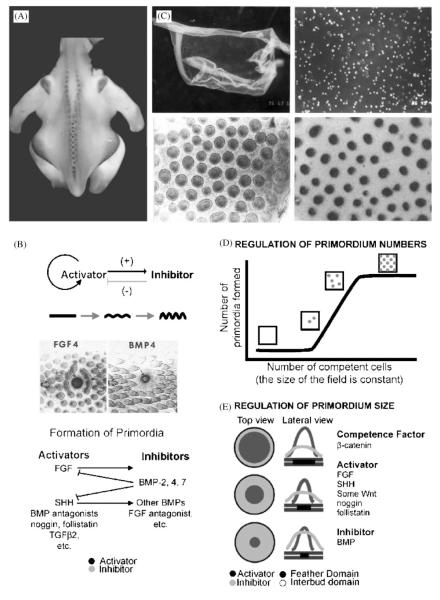
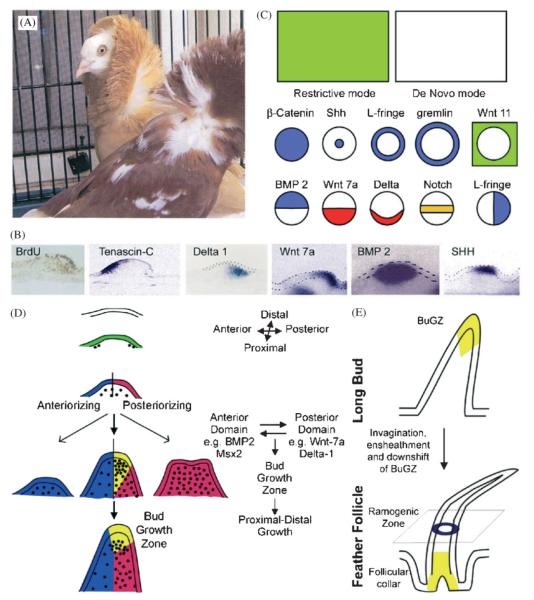
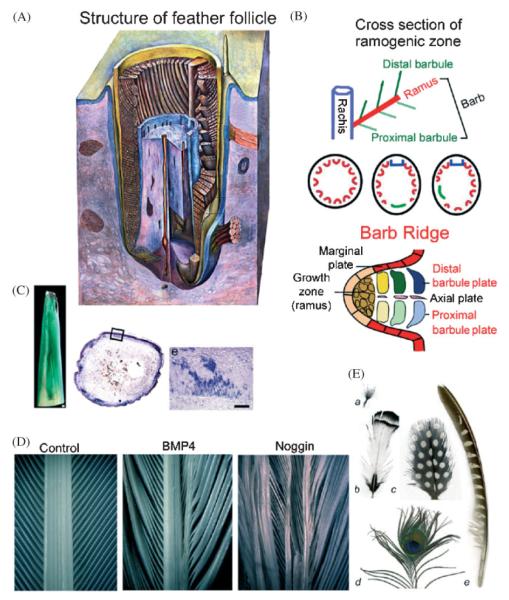
Darwin's theory describes the principles that are responsible for evolutionary change of organisms and their attributes. The actual mechanisms, however, need to be studied for each species and each organ separately. Here we have investigated the mechanisms underlying these principles in the avian feather. Feathers comprise one of the most complex and diverse epidermal organs as demonstrated by their shape, size, patterned arrangement and pigmentation. Variations can occur at several steps along each level of organization, leading to highly diverse forms and functions. Feathers develop gradually during ontogeny through a series of steps that may correspond to the evolutionary steps that were taken during the phylogeny from a reptilian ancestor to birds. These developmental steps include 1) the formation of feather tract fields on the skin surfaces; 2) periodic patterning of the individual feather primordia within the feather tract fields; 3) feather bud morphogenesis establishing anterio-posterior (along the cranio-caudal axis) and proximo-distal axes; 4) branching morphogenesis to create the rachis, barbs and barbules within a feather bud; and 5) gradual modulations of these basic morphological parameters within a single feather or across a feather tract. Thus, possibilities for variation in form and function of feathers occur at every developmental step. In this paper, principles guiding feather tract formation, distributions of individual feathers within the tracts and variations in feather forms are discussed at a cellular and molecular level.
Copyright 2003 Wiley-Liss, Inc.
Figures
References
-
- Bartels T, Meyer W, Neurand K. Skin and plumage changes in domestic birds. II. Plumage changes Tierarztl Prax. 1992;20:275–281. - PubMed
-
- Brantjes H, Barker N, van Es J, Clevers H. TCF: Lady Justice casting the final verdict on the outcome of Wnt signalling. Biol Chem. 2002;383:255–261. - PubMed
-
- Chen CWJ, Jung HS, Jiang TX, Chuong CM. Asymmetric expression of Notch, Serrate and Delta is associated with the anterior posterior axis of feather buds. Dev Biol. 1997;188:181–187. - PubMed
-
- Chen CW, Chuong CM. Avian integument provides multiple possibilities to analyse different phases of skin appendage morphogenesis. J Investig Dermato. Symp Proc. 1999;4:333–337. - PubMed
-
- Chen CWJ, Chuong CM. Dynamic expression of lunatic fringe during feather morphogenesis: a switch from medial-lateral to anterior-posterior asymmetry. Mech Dev. 2000;91:351–354. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
