

Tetrahydrobiopterin-dependent preservation of nitric oxide-mediated endothelial function in diabetes by targeted transgenic GTP-cyclohydrolase I overexpression
- PMID: 12952921
- PMCID: PMC182196
- DOI: 10.1172/JCI17786
Tetrahydrobiopterin-dependent preservation of nitric oxide-mediated endothelial function in diabetes by targeted transgenic GTP-cyclohydrolase I overexpression
Abstract
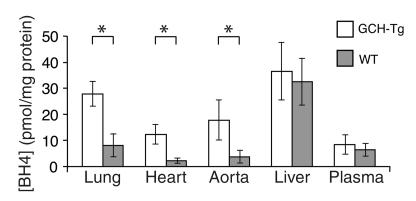
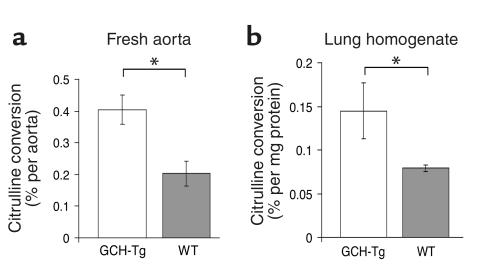
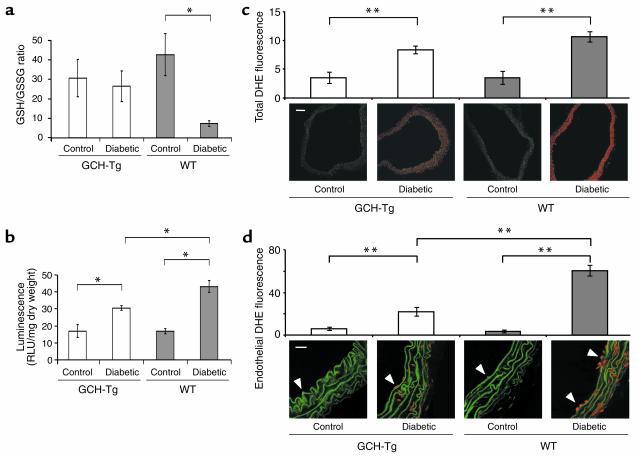
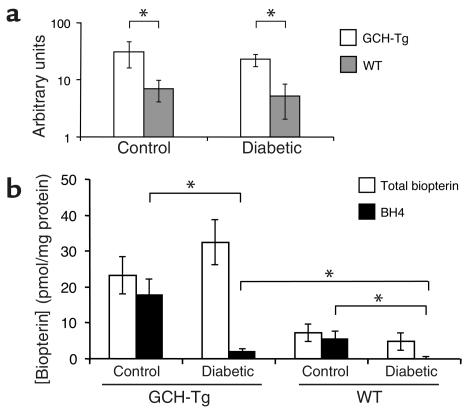
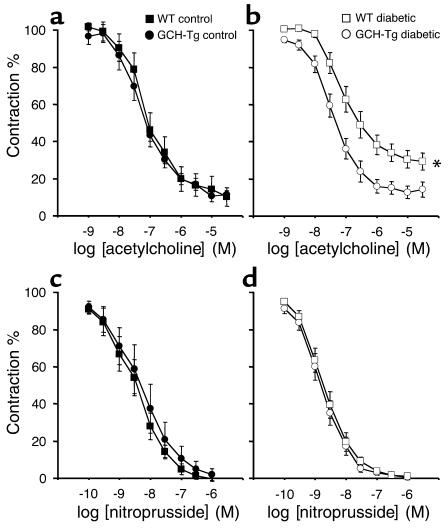
Increased production of reactive oxygen species and loss of endothelial NO bioactivity are key features of vascular disease states such as diabetes mellitus. Tetrahydrobiopterin (BH4) is a required cofactor for eNOS activity; pharmacologic studies suggest that BH4 may mediate some of the adverse effects of diabetes on eNOS function. We have now investigated the importance and mechanisms of BH4 availability in vivo using a novel transgenic mouse model with endothelial-targeted overexpression of the rate-limiting enzyme in BH4 synthesis, guanosine triphosphate-cyclohydrolase I (GTPCH). Transgenic (GCH-Tg) mice demonstrated selective augmentation of endothelial BH4 levels. In WT mice, induction of diabetes with streptozotocin (STZ) increased vascular oxidative stress, resulting in oxidative loss of BH4, forming BH2 and biopterin. Endothelial cell superoxide production in diabetes was increased, and NO-mediated endothelium-dependent vasodilatation was impaired. In diabetic GCH-Tg mice, superoxide production from the endothelium was markedly reduced compared with that of WT mice, endothelial BH4 levels were maintained despite some oxidative loss of BH4, and NO-mediated vasodilatation was preserved. These findings indicate that BH4 is an important mediator of eNOS regulation in diabetes and is a rational therapeutic target to restore NO-mediated endothelial function in diabetes and other vascular disease states.
Figures
References
-
- Cosentino F, Hishikawa K, Katusic ZS, Luscher TF. High glucose increases nitric oxide synthase expression and superoxide anion generation in human aortic endothelial cells. Circulation. 1997;96:25–28. - PubMed
-
- Cosentino F, Luscher TF. Tetrahydrobiopterin and endothelial nitric oxide synthase activity. Cardiovasc. Res. 1999;43:274–278. - PubMed
-
- Stuehr DJ. Mammalian nitric oxide synthases. . Biochim. Biophys. Acta. 1999; 1411:217–230. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
