

Local signaling by the EGF receptor
- PMID: 12952932
- PMCID: PMC2172819
- DOI: 10.1083/jcb.200303144
Local signaling by the EGF receptor
Abstract
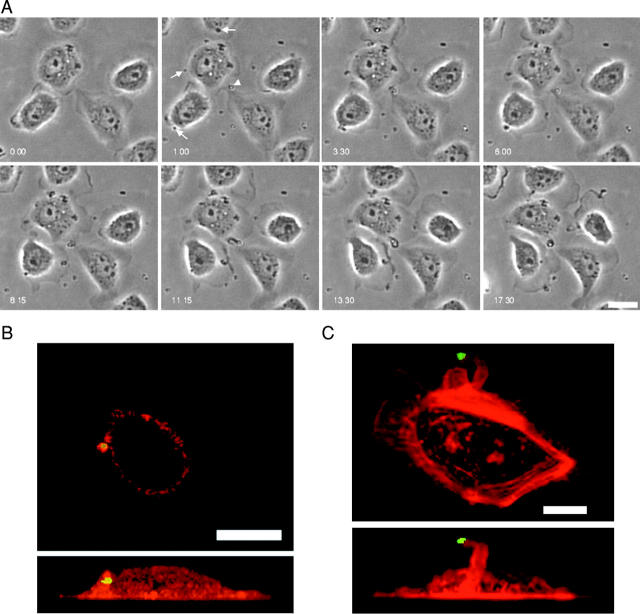
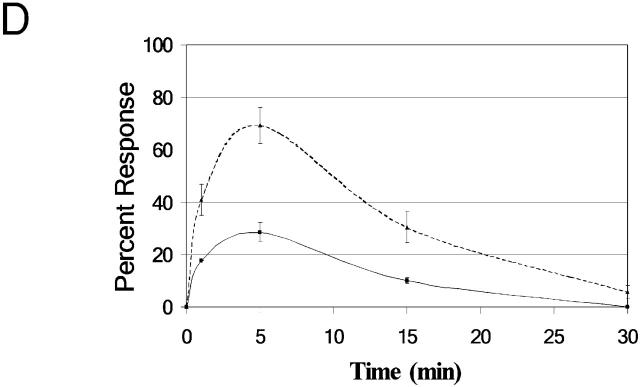
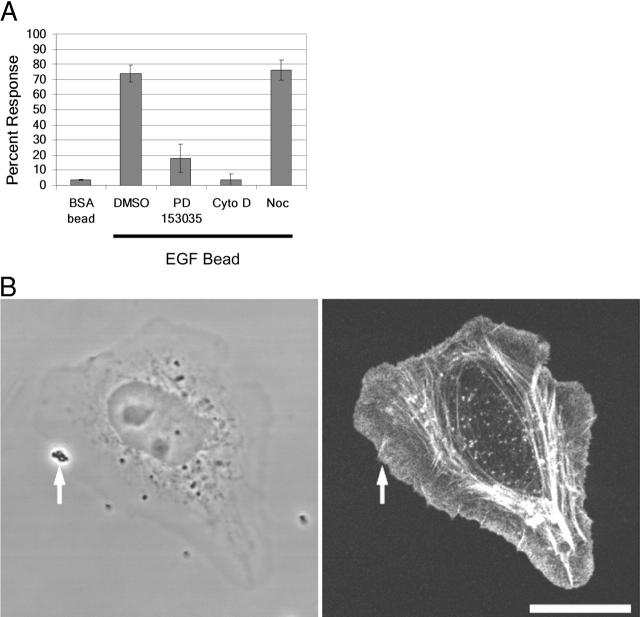
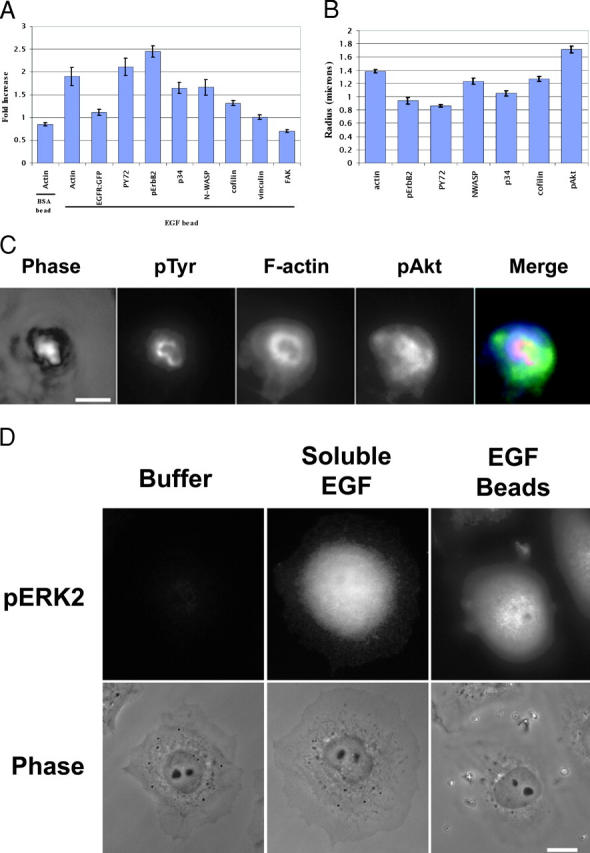
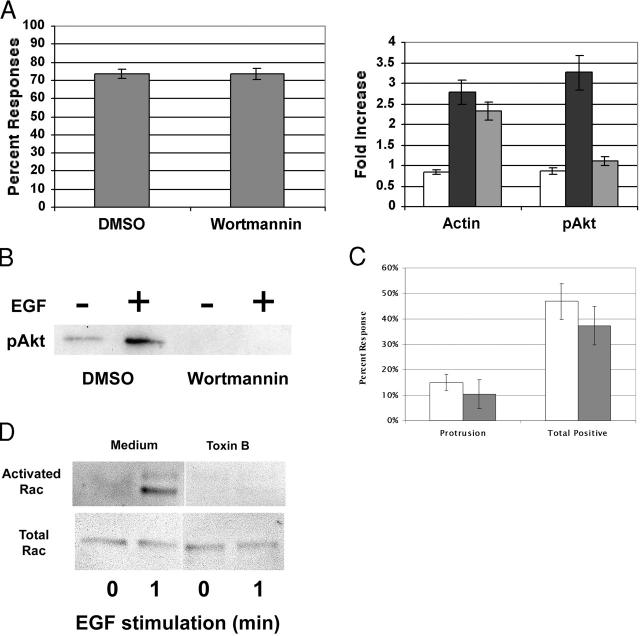
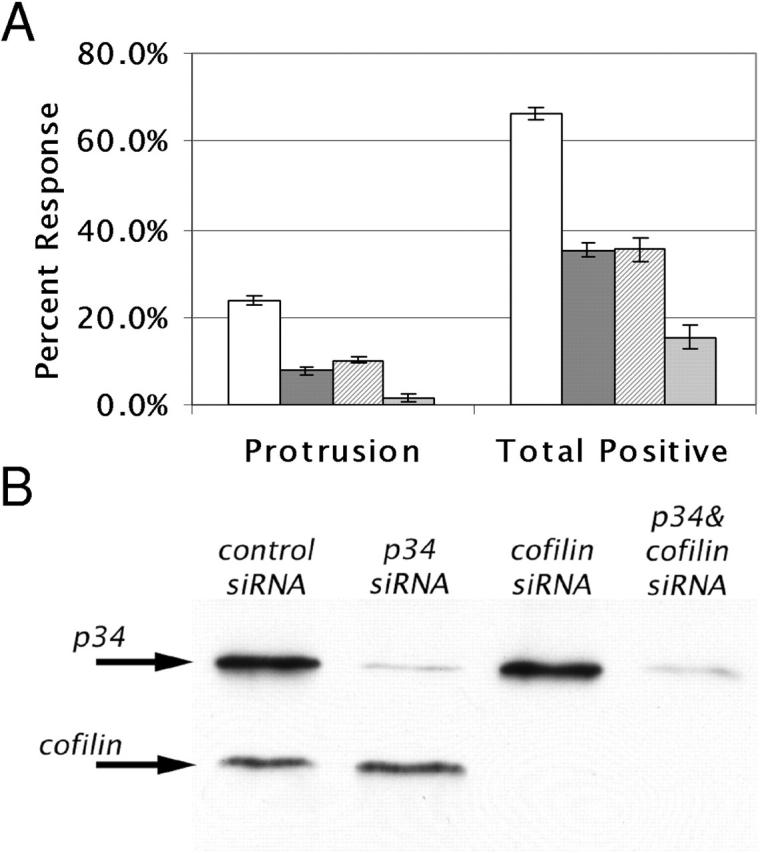
Differing spatial scales of signaling cascades are critical for cell orientation during chemotactic responses. We used biotin EGF bound to streptavidin-coupled magnetic beads to locally stimulate cells overexpressing the EGF receptor. We have found that EGF-induced actin polymerization remains localized even under conditions of receptor overexpression. Conversely, EGF-induced ERK activation spreads throughout the cell body after EGF bead stimulation. The localized actin polymerization is independent of PI3-kinase and rho protein activity and requires Arp2/3 complex and cofilin function. Thus, we find differing spatial scales of signaling from the EGF receptor, supporting models of chemotaxis that integrate short- and long-range signaling.
Figures
References
-
- Bailly, M., I. Ichetovkin, W. Grant, N. Zebda, L.M. Machesky, J.E. Segall, and J. Condeelis. 2001. The F-actin side binding activity of the Arp2/3 complex is essential for actin nucleation and lamellipod extension. Curr. Biol. 11:620–625. - PubMed
-
- Brock, R., and T.M. Jovin. 2001. Heterogeneity of signal transduction at the subcellular level: microsphere-based focal EGF receptor activation and stimulation of Shc translocation. J. Cell Sci. 114:2437–2447. - PubMed
-
- Condeelis, J.S., J.B. Wyckoff, M. Bailly, R. Pestell, D. Lawrence, J. Backer, and J.E. Segall. 2001. Lamellipodia in invasion. Semin. Cancer Biol. 11:119–128. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
