

A tale of two TUTases
- PMID: 12954983
- PMCID: PMC196853
- DOI: 10.1073/pnas.1833120100
A tale of two TUTases
Abstract
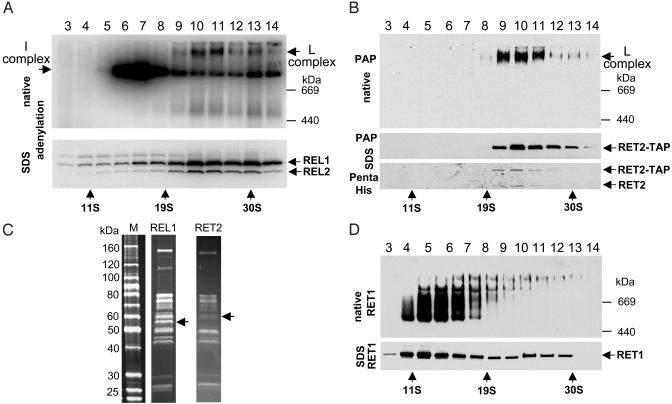
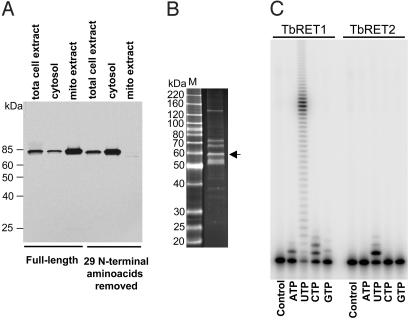
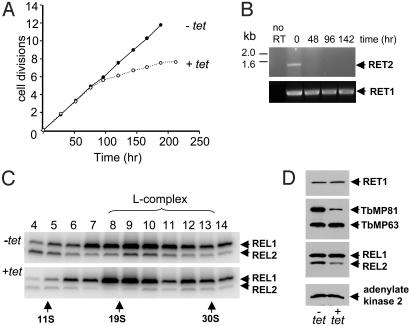
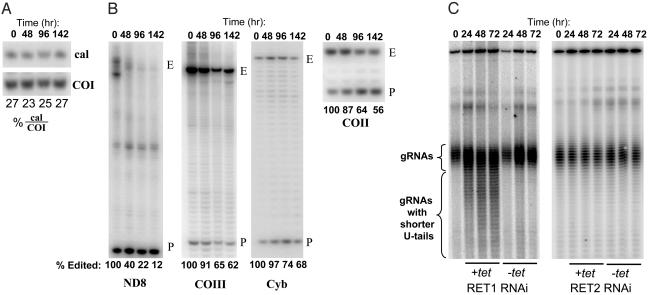
The insertion and deletion of U residues at specific sites in mRNAs in trypanosome mitochondria is thought to involve 3' terminal uridylyl transferase (TUTase) activity. TUTase activity is also required to create the nonencoded 3' oligo[U] tails of the transacting guide RNAs (gRNAs). We have described two TUTases, RET1 (RNA editing TUTase 1) and RET2 (RNA editing TUTase 2) as components of different editing complexes. Tandem affinity purification-tagged Trypanosoma brucei RET2 (TbRET2) was expressed and localized to the cytosol in Leishmania tarentolae cells by removing the mitochondrial signal sequence. Double-affinity isolation yielded tagged TbRET2, together with a few additional proteins. This material exhibits a U-specific transferase activity in which a single U is added to the 3' end of a single-stranded RNA, thereby confirming that RET2 is a 3' TUTase. We also found that RNA interference of RET2 expression in T. brucei inhibits in vitro U-insertion editing and has no effect on the length of the 3' oligo[U] tails of the gRNAs, whereas down-regulation of RET1 has a minor effect on in vitro U-insertion editing, but produces a decrease in the average length of the oligo[U] tails. This finding suggests that RET2 is responsible for U-insertions at editing sites and RET1 is involved in gRNA 3' end maturation, which is essential for creating functional gRNAs. From these results we have functionally relabeled the previously described TUT-II complex containing RET1 as the guide RNA processing complex.
Figures
Comment in
-
Two distinct roles for terminal uridylyl transferases in RNA editing.Proc Natl Acad Sci U S A. 2003 Sep 16;100(19):10583-4. doi: 10.1073/pnas.2035062100. Epub 2003 Sep 8. Proc Natl Acad Sci U S A. 2003. PMID: 12963809 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
