

The angiotensin II type I receptor-associated protein, ATRAP, is a transmembrane protein and a modulator of angiotensin II signaling
- PMID: 12960423
- PMCID: PMC284805
- DOI: 10.1091/mbc.e03-06-0383
The angiotensin II type I receptor-associated protein, ATRAP, is a transmembrane protein and a modulator of angiotensin II signaling
Abstract
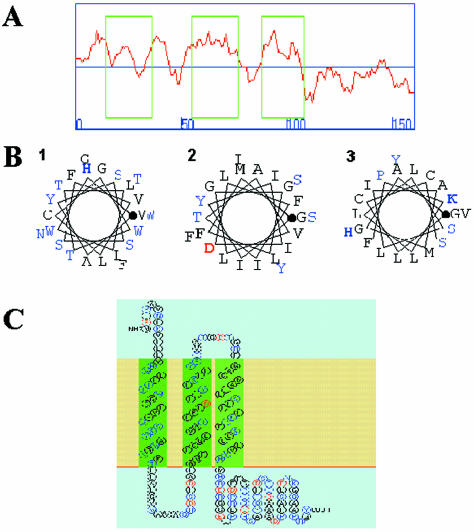
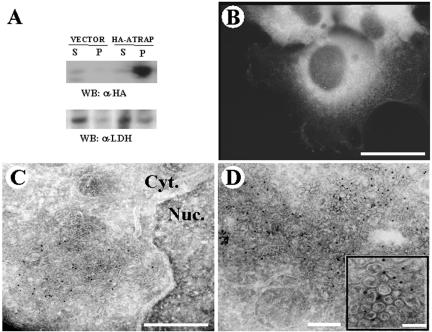
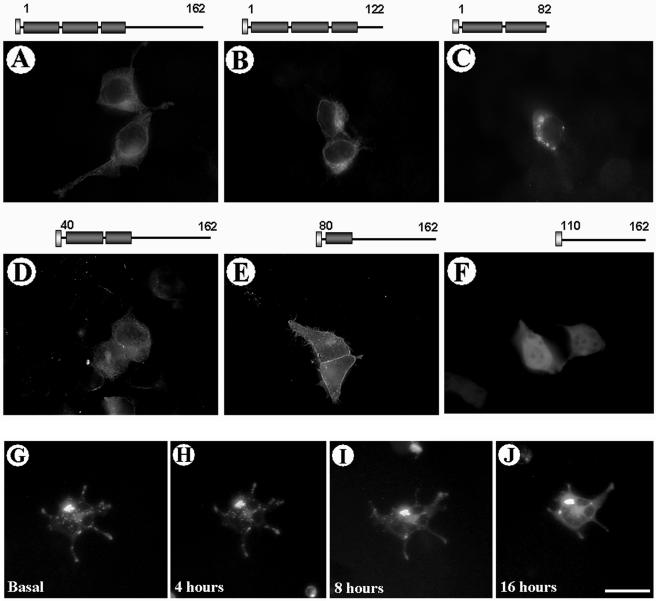
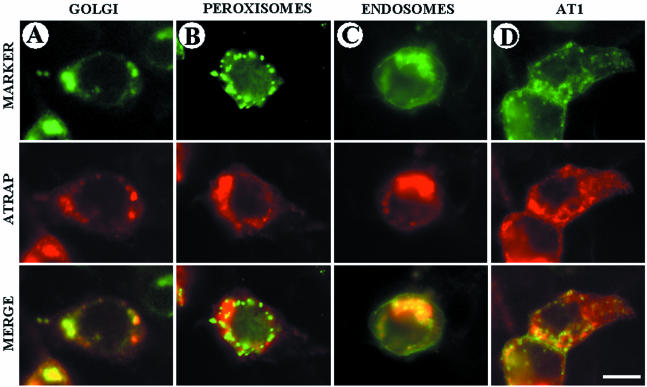
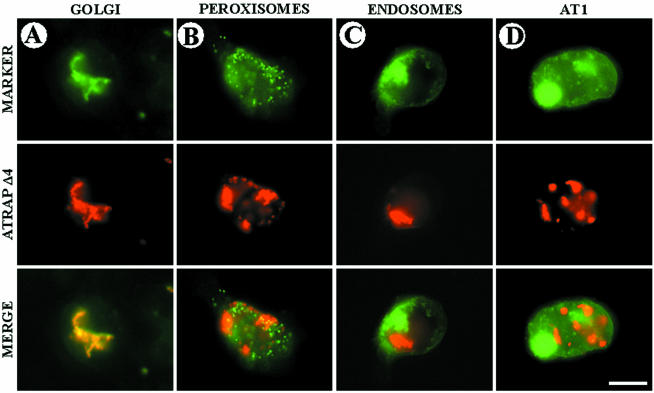
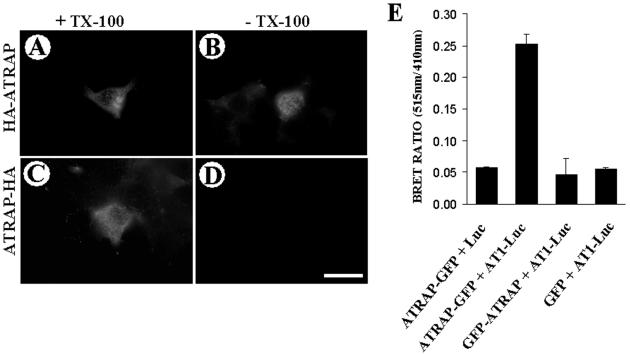
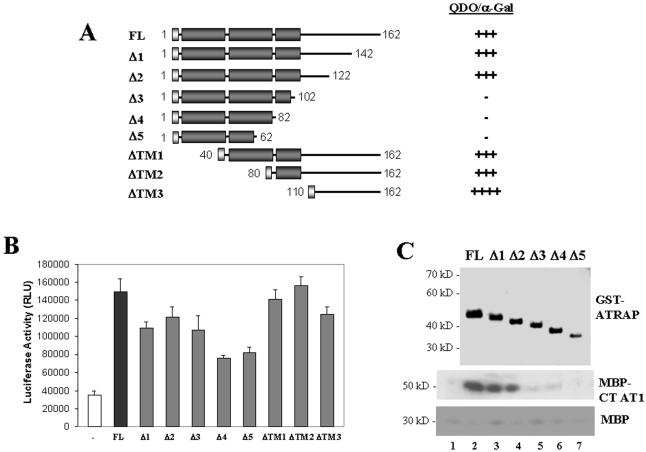
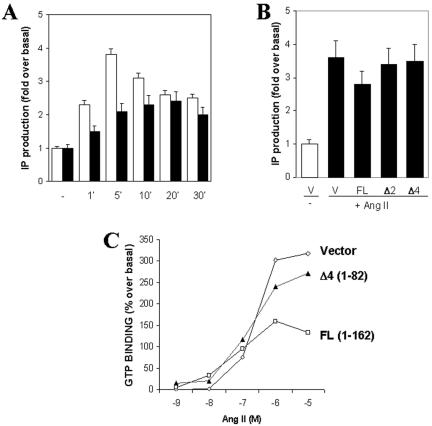
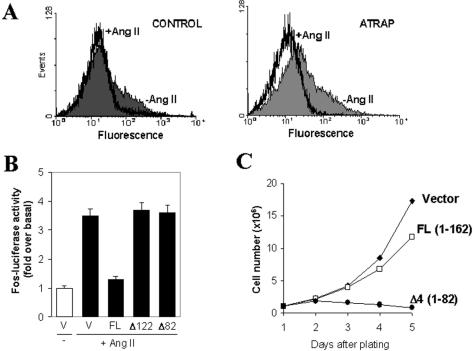
Our group identified angiotensin II type 1 (AT1) receptor-associated protein (ATRAP) in a yeast two-hybrid screen for proteins that bind to the carboxyl-terminal cytoplasmic domain of the AT1. In this work, we characterize ATRAP as a transmembrane protein localized in intracellular trafficking vesicles and plasma membrane that functions as a modulator of angiotensin II-induced signal transduction. ATRAP contains three hydrophobic domains at the amino-terminal end of the protein, encompassing the amino acid residues 14-36, 55-77, and 88-108 and a hydrophilic cytoplasmic carboxyl-terminal tail from residues 109-161. Endogenous and transfected ATRAP cDNA shows a particulate distribution; electron microscopy reveals the presence of ATRAP in prominent perinuclear vesicular membranes; and colocalization analysis by immunofluorescence shows that ATRAP colocalizes in an intracellular vesicular compartment corresponding to endoplasmic reticulum, Golgi, and endocytic vesicles. Real-time tracking of ATRAP vesicles shows constitutive translocation toward the plasma membrane. Using epitope-tagged forms of ATRAP at either the amino or carboxyl end of the molecule, we determined the orientation of the amino end as being outside the cell. Mutant forms of ATRAP lacking the carboxyl end are unable to bind to the AT1 receptor, leading to the formation of prominent perinuclear vesicle clusters. Functional analysis of the effects of ATRAP on angiotensin II-induced AT1 receptor signaling reveals a moderate decrease in the generation of inositol lipids, a marked decrease in the angiotensin II-stimulated transcriptional activity of the c-fos promoter luciferase reporter gene, and a decrease in cell proliferation.
Figures
Similar articles
-
Identification of calcium-modulating cyclophilin ligand (CAML) as transducer of angiotensin II-mediated nuclear factor of activated T cells (NFAT) activation.J Biol Chem. 2005 Apr 1;280(13):12536-41. doi: 10.1074/jbc.M500296200. Epub 2005 Jan 24. J Biol Chem. 2005. PMID: 15668245
-
Cloning and characterization of ATRAP, a novel protein that interacts with the angiotensin II type 1 receptor.J Biol Chem. 1999 Jun 11;274(24):17058-62. doi: 10.1074/jbc.274.24.17058. J Biol Chem. 1999. PMID: 10358057
-
ATRAP, novel AT1 receptor associated protein, enhances internalization of AT1 receptor and inhibits vascular smooth muscle cell growth.Biochem Biophys Res Commun. 2000 Dec 29;279(3):938-41. doi: 10.1006/bbrc.2000.4055. Biochem Biophys Res Commun. 2000. PMID: 11162453
-
The role of angiotensin AT1 receptor-associated protein in renin-angiotensin system regulation and function.Curr Hypertens Rep. 2007 Apr;9(2):121-7. doi: 10.1007/s11906-007-0022-6. Curr Hypertens Rep. 2007. PMID: 17442223 Review.
-
Angiotensin II Type 1 Receptor Binding Molecule ATRAP as a Possible Modulator of Renal Sodium Handling and Blood Pressure in Pathophysiology.Curr Med Chem. 2015;22(28):3210-6. doi: 10.2174/0929867322666150821095036. Curr Med Chem. 2015. PMID: 26295465 Review.
Cited by
-
From cell surface to nucleus: Mas transportation in hypertension.Cardiovasc Res. 2020 Oct 1;116(12):1929-1931. doi: 10.1093/cvr/cvaa087. Cardiovasc Res. 2020. PMID: 32246824 Free PMC article. No abstract available.
-
Effects of ATRAP in Renal Proximal Tubules on Angiotensin-Dependent Hypertension.J Am Heart Assoc. 2019 Apr 16;8(8):e012395. doi: 10.1161/JAHA.119.012395. J Am Heart Assoc. 2019. PMID: 30977419 Free PMC article.
-
S-adenosylmethionine upregulates the angiotensin receptor-binding protein ATRAP via the methylation of HuR in NAFLD.Cell Death Dis. 2021 Mar 22;12(4):306. doi: 10.1038/s41419-021-03591-1. Cell Death Dis. 2021. PMID: 33753727 Free PMC article.
-
The AT1/AT2 Receptor Equilibrium Is a Cornerstone of the Regulation of the Renin Angiotensin System beyond the Cardiovascular System.Molecules. 2023 Jul 18;28(14):5481. doi: 10.3390/molecules28145481. Molecules. 2023. PMID: 37513355 Free PMC article. Review.
-
Deletion of the angiotensin II type 1 receptor-associated protein enhances renal sodium reabsorption and exacerbates angiotensin II-mediated hypertension.Kidney Int. 2014 Sep;86(3):570-81. doi: 10.1038/ki.2014.95. Epub 2014 Apr 2. Kidney Int. 2014. PMID: 24694992 Free PMC article.
References
-
- Allen, A.M., Zhuo, J., and Mendelsohn, F.A. (2000). Localization and function of angiotensin AT1 receptors. Am. J. Hypertens. 13, 31S-38S. - PubMed
-
- Audoly, L.P., Oliverio, M.I., and Coffman, T.M. (2000). Insights into the functions of type 1 (AT1) angiotensin II receptors provided by gene targeting. Trends Endocrinol. Metab. 11, 263-269. - PubMed
-
- Blume, A., Herdegen, T., and Unger, T. (1999). Angiotensin peptides and inducible transcription factors. J. Mol. Med. 77, 339-357. - PubMed
-
- Brzostowski, J.A., and Kimmel, A.R. (2001). Signaling at zero G: G-protein-independent functions for 7-TM receptors. Trends Biochem. Sci. 26, 291-297. - PubMed
-
- Conchon, S., Barrault, M.B., Miserey, S., Corvol, P., and Clauser, E. (1997). The C-terminal third intracellular loop of the rat AT1 angiotensin II receptor plays a key role in G-protein coupling specificity and transduction of the mitogenic signal. J. Biol. Chem. 272, 25566-25572. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials