

Allosteric switching by mutually exclusive folding of protein domains
- PMID: 12963365
- PMCID: PMC3145375
- DOI: 10.1016/s0022-2836(03)00925-2
Allosteric switching by mutually exclusive folding of protein domains
Abstract
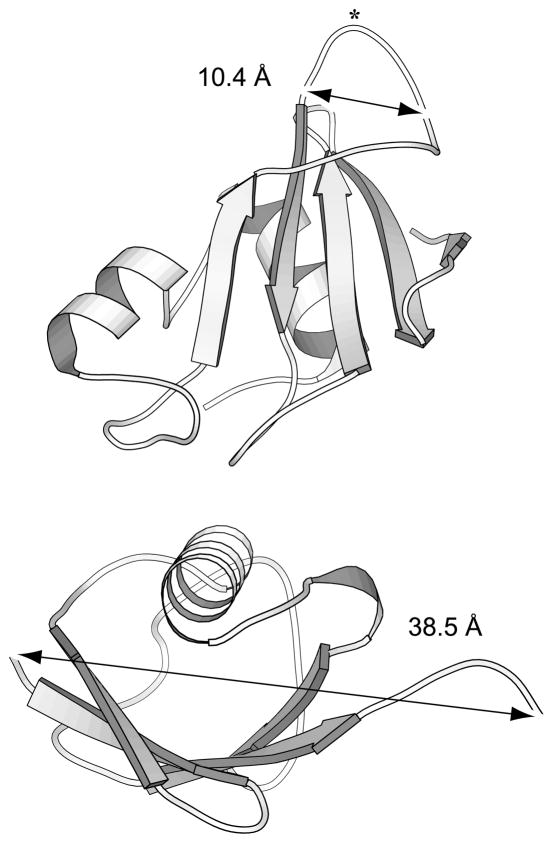
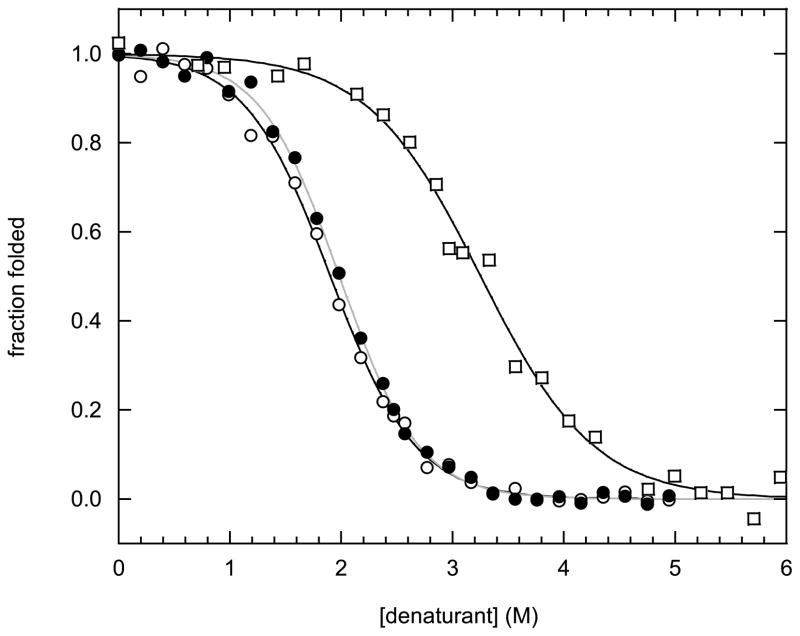
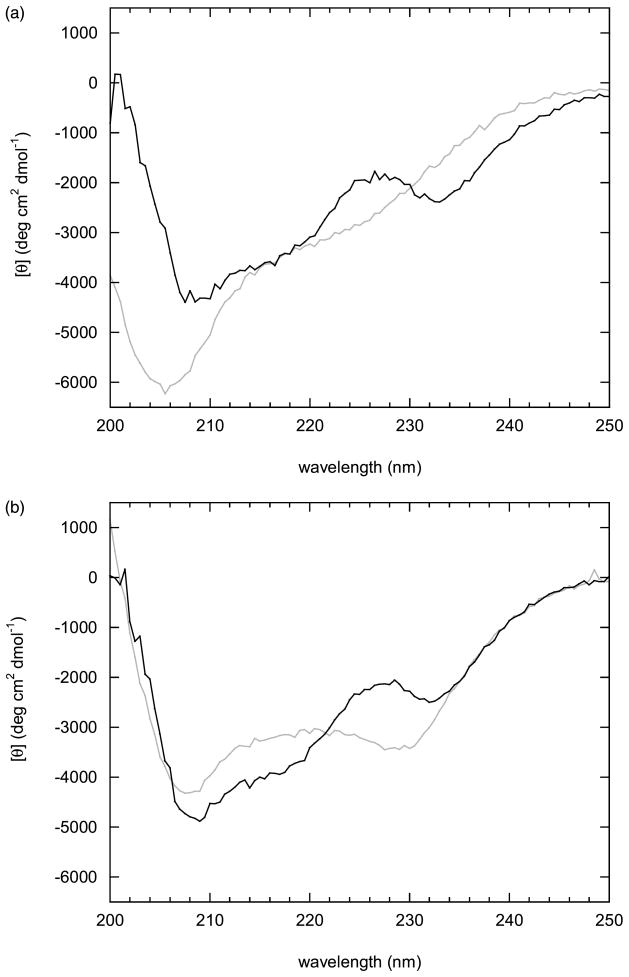
Many proteins are built from structurally and functionally distinct domains. A major goal is to understand how conformational change transmits information between domains in order to achieve biological activity. A two-domain, bi-functional fusion protein has been designed so that the mechanical stress imposed by the folded structure of one subunit causes the other subunit to unfold, and vice versa. The construct consists of ubiquitin inserted into a surface loop of barnase. The distance between the amino and carboxyl ends of ubiquitin is much greater than the distance between the termini of the barnase loop. This topological constraint causes the two domains to engage in a thermodynamic tug-of-war in which only one can exist in its folded state at any given time. This conformational equilibrium, which is cooperative, reversible, and controllable by ligand binding, serves as a model for the coupled binding and folding mechanism widely used to mediate protein-protein interactions and cellular signaling processes. The position of the equilibrium can be adjusted by temperature or ligand binding and is monitored in vivo by cell death. This design forms the basis for a new class of cytotoxic proteins that can be activated by cell-specific effector molecules, and can thus target particular cell types for destruction.
Figures


References
-
- Dyson HJ, Wright PE. Coupling of folding and binding for unstructured proteins. Curr Opin Struct Biol. 2002;12:54–60. - PubMed
-
- Luque I, Leavitt SA, Friere E. The linkage between protein folding and functional cooperativity: two sides of the same coin. Annu Rev Biophys Biomol Struct. 2002;31:235–256. - PubMed
-
- Botuyan MV, Mer G, Yi GS, Koth CM, Case DA, Edwards AM, et al. Solution structure and dynamics of yeast elongin C in complex with a von-Hippel-Lindau peptide. J Mol Biol. 2001;312:177–186. - PubMed
-
- Kim AS, Kakalis LT, Abdul-Manan N, Liu GA, Rosen MK. Autoinhibition and activation mechanisms of the Wiskott–Aldrich syndrome protein. Nature. 2000;404:151–158. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
