

Modulation of synaptic transmission by the BCL-2 family protein BCL-xL
- PMID: 12968005
- PMCID: PMC6740692
- DOI: 10.1523/JNEUROSCI.23-23-08423.2003
Modulation of synaptic transmission by the BCL-2 family protein BCL-xL
Abstract
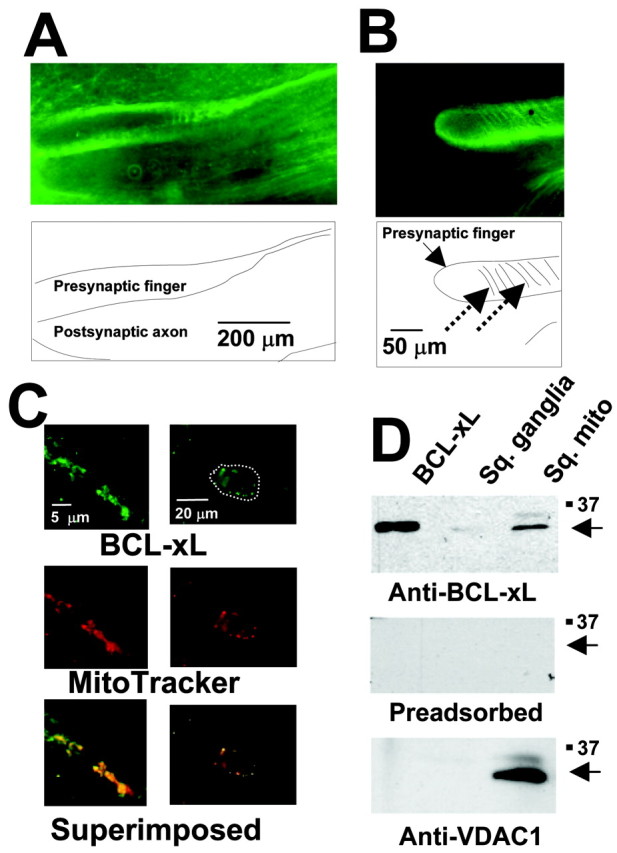
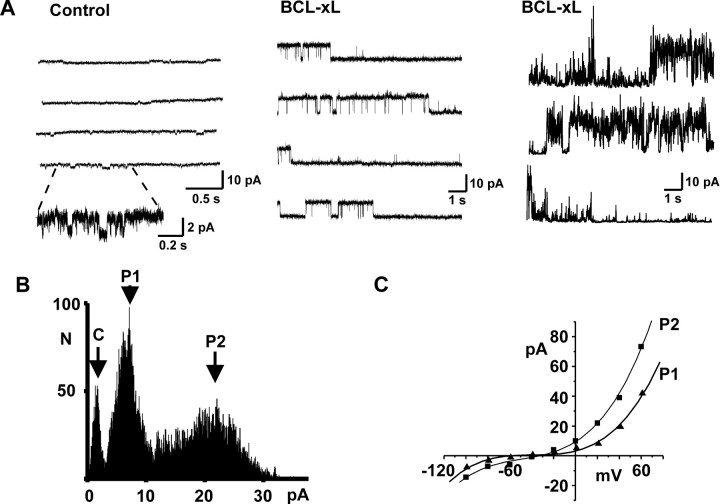
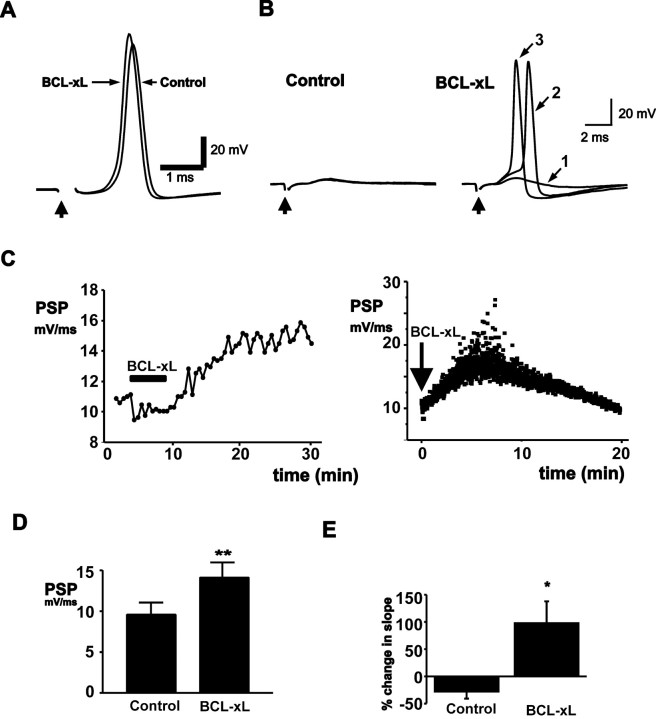
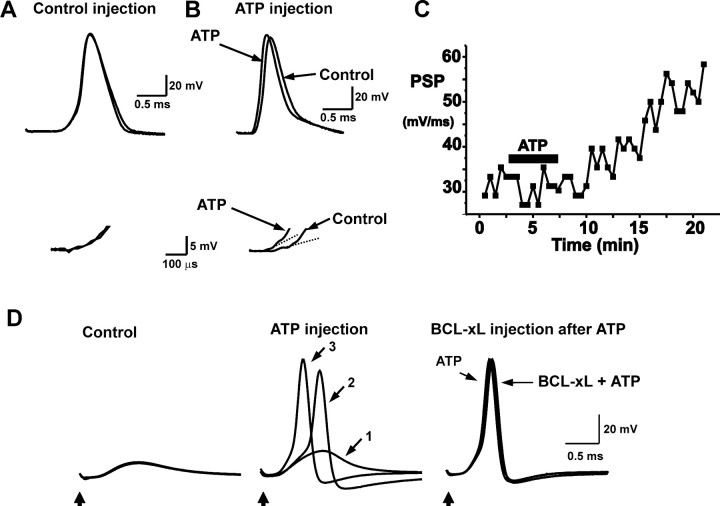
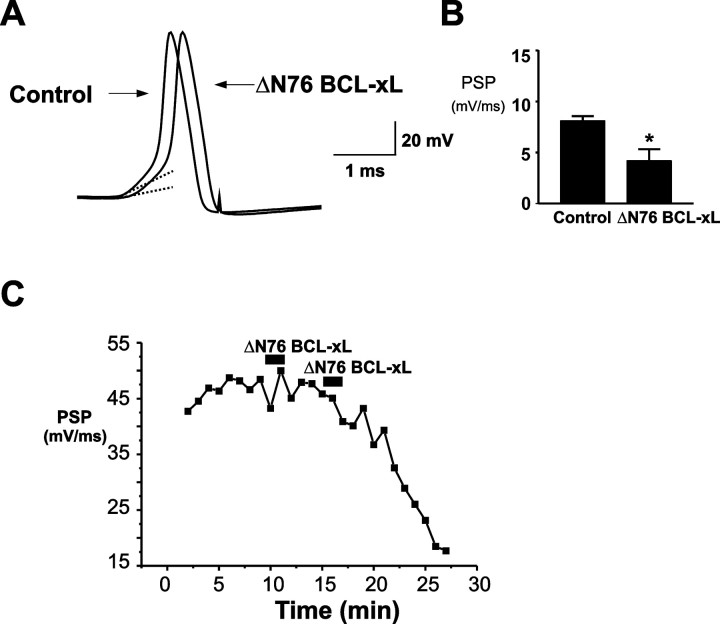
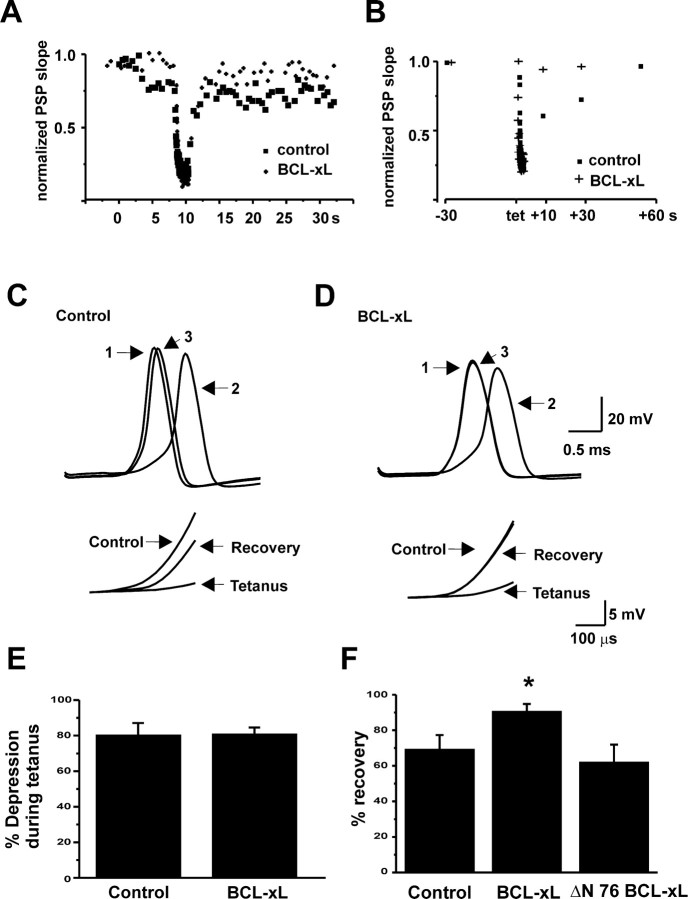
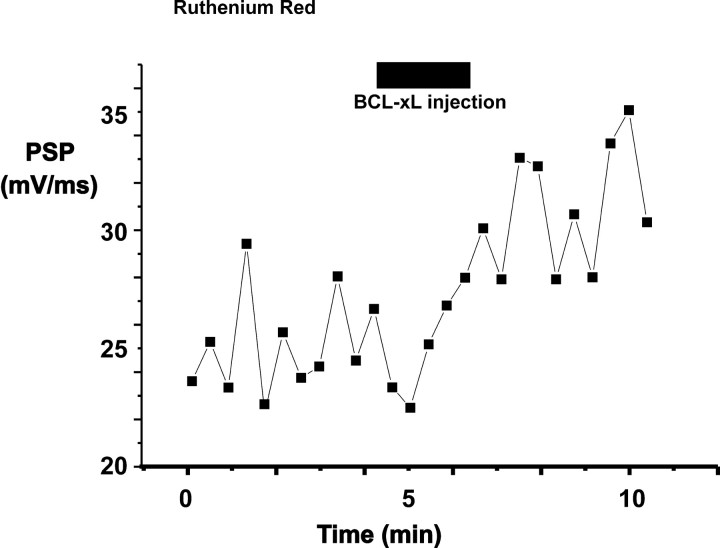
BCL-2 family proteins are known to regulate cell death during development by influencing the permeability of mitochondrial membranes. The anti-apoptotic BCL-2 family protein BCL-xL is highly expressed in the adult brain and localizes to mitochondria in the presynaptic terminal of the adult squid stellate ganglion. Application of recombinant BCL-xL through a patch pipette to mitochondria inside the giant presynaptic terminal triggered multiconductance channel activity in mitochondrial membranes. Furthermore, injection of full-length BCL-xL protein into the presynaptic terminal enhanced postsynaptic responses and enhanced the rate of recovery from synaptic depression, whereas a recombinant pro-apoptotic cleavage product of BCL-xL attenuated postsynaptic responses. The effect of BCL-xL on synaptic responses persisted in the presence of a blocker of mitochondrial calcium uptake and was mimicked by injection of ATP into the terminal. These studies indicate that the permeability of outer mitochondrial membranes influences synaptic transmission, and they raise the possibility that modulation of mitochondrial conductance by BCL-2 family proteins affects synaptic stability.
Figures
References
-
- Antonsson B, Conti F, Ciavatta A, Montessuit S, Lewis S, Martinou I, Bernasconi L, Bernard A, Mermod JJ, Mazzei G, Maundrell K, Gambale F, Sadoul R, Martinou JC ( 1997) Inhibition of Bax channel-forming activity by Bcl-2. Science 277 : 370-372. - PubMed
-
- Augustine GJ ( 2001) How does calcium trigger neurotransmitter release? Curr Opin Neurobiol 11 : 320-326. - PubMed
-
- Babcock DF, Hille B ( 1998) Mitochondrial oversight of cellular Ca2+ signaling. Curr Opin Neurobiol 8 : 398-404. - PubMed
-
- Basanez G, Zhang J, Chau BN, Maksaev GI, Frolov VA, Brandt TA, Burch J, Hardwick JM, Zimmerberg J ( 2001) Pro-apoptotic cleavage products of Bcl-xL form cytochrome c-conducting pores in pure lipid membranes. J Biol Chem 276 : 31083-31091. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials