

Programmed translational -1 frameshifting on hexanucleotide motifs and the wobble properties of tRNAs
- PMID: 12970189
- PMCID: PMC212731
- DOI: 10.1093/emboj/cdg465
Programmed translational -1 frameshifting on hexanucleotide motifs and the wobble properties of tRNAs
Abstract
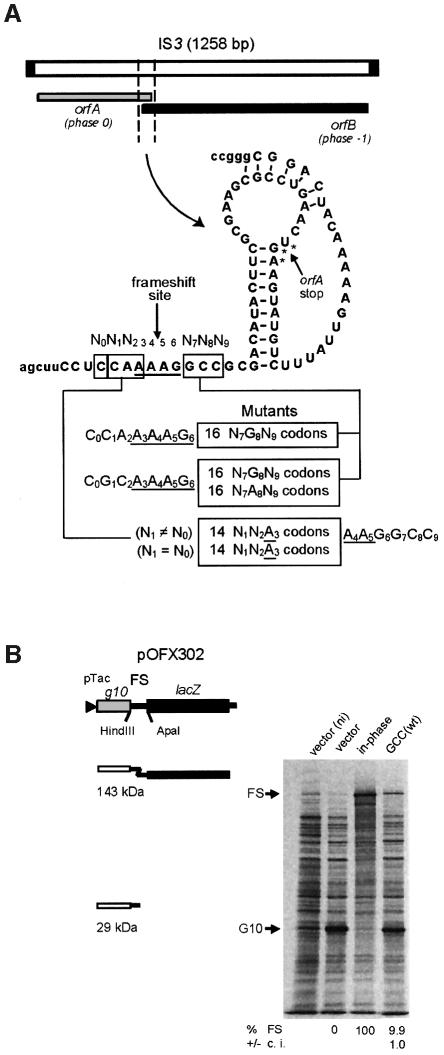
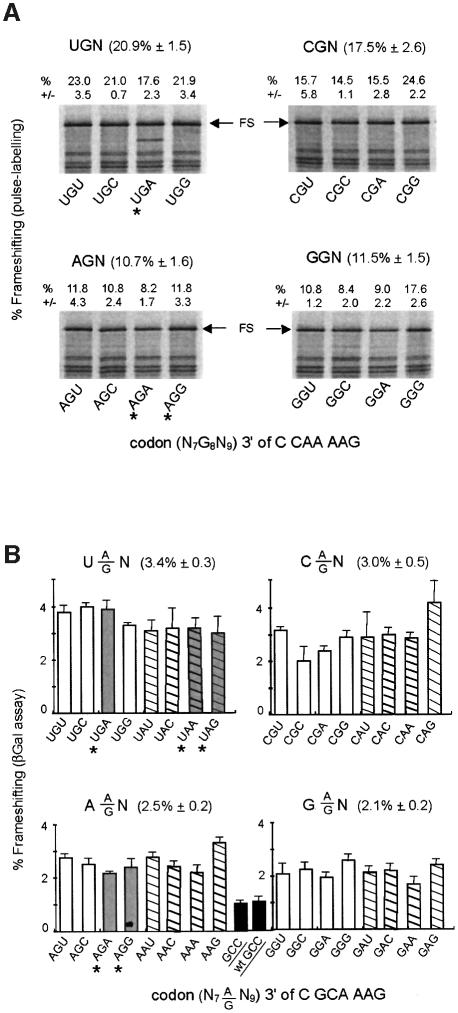
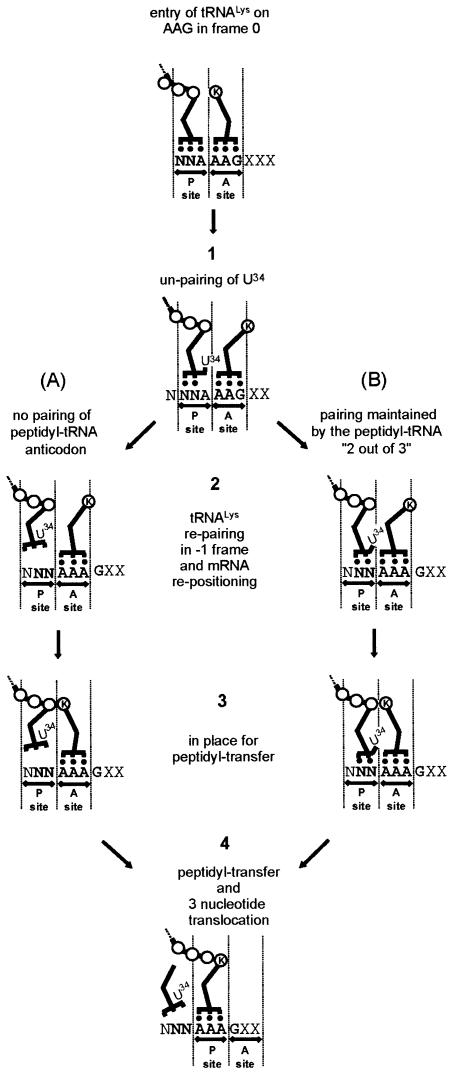
Programmed -1 ribosomal frameshifting, involving tRNA re-pairing from an AAG codon to an AAA codon, has been reported to occur at the sequences CGA AAG and CAA AAG. In this study, using the recoding region of insertion sequence IS3, we have investigated the influence on frameshifting in Escherichia coli of the first codon of this type of motif by changing it to all other NNA codons. Two classes of NNA codons were distinguished, depending on whether they favor or limit frameshifting. Their degree of shiftiness is correlated with wobble propensity, and base 34 modification, of their decoding tRNAs. A more flexible anticodon loop very likely makes the tRNAs with extended wobble more prone to liberate the third codon base, A, for re-pairing of tRNALys in the -1 frame.
Figures


References
-
- Atkins J.F. and Gesteland,R.F. (1995) Discontinuous triplet decoding with or without re-pairing by peptidyl tRNA. In Söll,D. and RajBhandary,U.L. (eds), tRNA: Structure, Biosynthesis and Function. ASM Press, Washington, DC, pp. 471–490.
-
- Atkins J.F., Herr,A., Massire,C., O’Connor,M., Ivanov,I. and Gesteland,R.F. (2000) Poking a hole in the sanctity of the triplet code: inferences for framing. In Garrett,R.A., Douthwaite,S.R., Liljas,A., Matheson,A.T., Moore,P.B. and Noller,H.F. (eds), The Ribosome: Structure, Function, Antibiotics and Cellular Interactions. ASM Press, Washington, DC, pp. 369–383.
-
- Atkins J.F. et al. (2001) Over-riding standard decoding: implication of recoding for ribosome function and enrichment of gene expression. In Cold Spring Harbor Symposium on Quantitative Biology, Vol. LXVI, The Ribosome. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, pp. 217–232. - PubMed
-
- Björk G.R. (1995) Biosynthesis and function of modified nucleosides in tRNA. In Söll,D. and RajBhandary,U.L. (eds), tRNA: Structure, Biosynthesis and Function. ASM Press, Washington, DC, pp. 165–205.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
