

Characterization of RNA elements that regulate gag-pol ribosomal frameshifting in equine infectious anemia virus
- PMID: 12970412
- PMCID: PMC228510
- DOI: 10.1128/jvi.77.19.10280-10287.2003
Characterization of RNA elements that regulate gag-pol ribosomal frameshifting in equine infectious anemia virus
Abstract
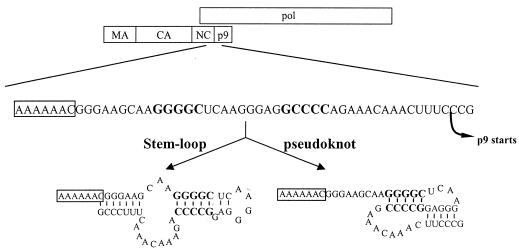
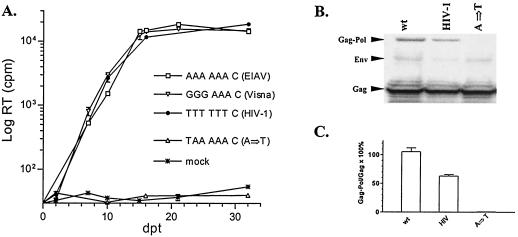
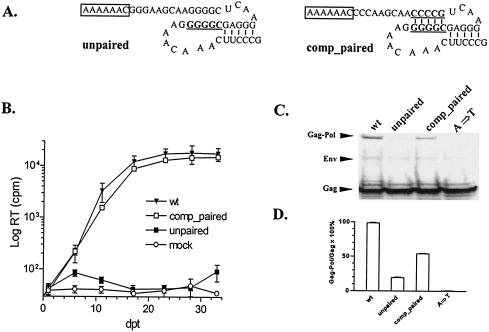
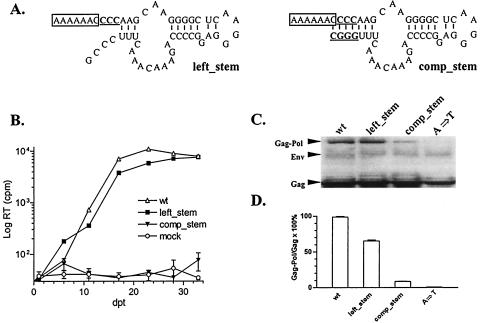
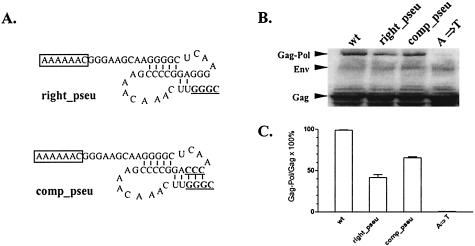
Synthesis of Gag-Pol polyproteins of retroviruses requires ribosomes to shift translational reading frame once or twice in a -1 direction to read through the stop codon in the gag reading frame. It is generally believed that a slippery sequence and a downstream RNA structure are required for the programmed -1 ribosomal frameshifting. However, the mechanism regulating the Gag-Pol frameshifting remains poorly understood. In this report, we have defined specific mRNA elements required for sufficient ribosomal frameshifting in equine anemia infectious virus (EIAV) by using full-length provirus replication and Gag/Gag-Pol expression systems. The results of these studies revealed that frameshifting efficiency and viral replication were dependent on a characteristic slippery sequence, a five-base-paired GC stretch, and a pseudoknot structure. Heterologous slippery sequences from human immunodeficiency virus type 1 and visna virus were able to substitute for the EIAV slippery sequence in supporting EIAV replication. Disruption of the GC-paired stretch abolished the frameshifting required for viral replication, and disruption of the pseudoknot reduced the frameshifting efficiency by 60%. Our data indicated that maintenance of the essential RNA signals (slippery sequences and structural elements) in this region of the genomic mRNA was critical for sufficient ribosomal frameshifting and EIAV replication, while concomitant alterations in the amino acids translated from the same region of the mRNA could be tolerated during replication. The data further indicated that proviral mutations that reduced frameshifting efficiency by as much as 50% continued to sustain viral replication and that greater reductions in frameshifting efficiency lead to replication defects. These studies define for the first time the RNA sequence and structural determinants of Gag-Pol frameshifting necessary for EIAV replication, reveal novel aspects relative to frameshifting elements described for other retroviruses, and provide new genetic determinants that can be evaluated as potential antiviral targets.
Figures
Similar articles
-
Secondary structure and mutational analysis of the ribosomal frameshift signal of rous sarcoma virus.J Mol Biol. 1998 Nov 27;284(2):205-25. doi: 10.1006/jmbi.1998.2186. J Mol Biol. 1998. PMID: 9813113 Free PMC article.
-
Modulation of HIV-1 Gag/Gag-Pol frameshifting by tRNA abundance.Nucleic Acids Res. 2019 Jun 4;47(10):5210-5222. doi: 10.1093/nar/gkz202. Nucleic Acids Res. 2019. PMID: 30968122 Free PMC article.
-
Identification and analysis of the gag-pol ribosomal frameshift site of feline immunodeficiency virus.Virology. 1992 Feb;186(2):389-97. doi: 10.1016/0042-6822(92)90004-9. Virology. 1992. PMID: 1310175 Free PMC article.
-
A review on architecture of the gag-pol ribosomal frameshifting RNA in human immunodeficiency virus: a variability survey of virus genotypes.J Biomol Struct Dyn. 2017 Jun;35(8):1629-1653. doi: 10.1080/07391102.2016.1194231. Epub 2016 Aug 2. J Biomol Struct Dyn. 2017. PMID: 27485859 Review.
-
Structure, stability and function of RNA pseudoknots involved in stimulating ribosomal frameshifting.J Mol Biol. 2000 Apr 28;298(2):167-85. doi: 10.1006/jmbi.2000.3668. J Mol Biol. 2000. PMID: 10764589 Free PMC article. Review.
Cited by
-
A Functional Interplay between Human Immunodeficiency Virus Type 1 Protease Residues 77 and 93 Involved in Differential Regulation of Precursor Autoprocessing and Mature Protease Activity.PLoS One. 2015 Apr 20;10(4):e0123561. doi: 10.1371/journal.pone.0123561. eCollection 2015. PLoS One. 2015. PMID: 25893662 Free PMC article.
-
Modulation of human immunodeficiency virus type 1 protease autoprocessing by charge properties of surface residue 69.J Virol. 2009 Aug;83(15):7789-93. doi: 10.1128/JVI.00473-09. Epub 2009 May 20. J Virol. 2009. PMID: 19457992 Free PMC article.
-
Targeting HIV-1 Protease Autoprocessing for High-throughput Drug Discovery and Drug Resistance Assessment.Sci Rep. 2019 Jan 22;9(1):301. doi: 10.1038/s41598-018-36730-4. Sci Rep. 2019. PMID: 30670786 Free PMC article.
-
Understanding HIV-1 protease autoprocessing for novel therapeutic development.Future Med Chem. 2013 Jul;5(11):1215-29. doi: 10.4155/fmc.13.89. Future Med Chem. 2013. PMID: 23859204 Free PMC article.
-
Ribosomal frameshifting and transcriptional slippage: From genetic steganography and cryptography to adventitious use.Nucleic Acids Res. 2016 Sep 6;44(15):7007-78. doi: 10.1093/nar/gkw530. Epub 2016 Jul 19. Nucleic Acids Res. 2016. PMID: 27436286 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
