

Purification, identification, and biochemical characterization of a host-encoded cysteine protease that cleaves a leishmaniavirus gag-pol polyprotein
- PMID: 12970430
- PMCID: PMC228495
- DOI: 10.1128/jvi.77.19.10448-10455.2003
Purification, identification, and biochemical characterization of a host-encoded cysteine protease that cleaves a leishmaniavirus gag-pol polyprotein
Abstract
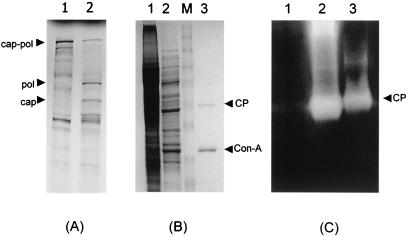
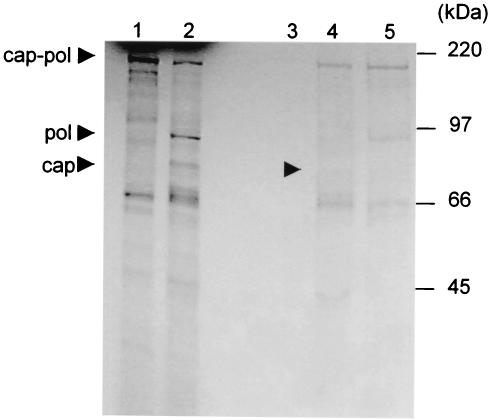
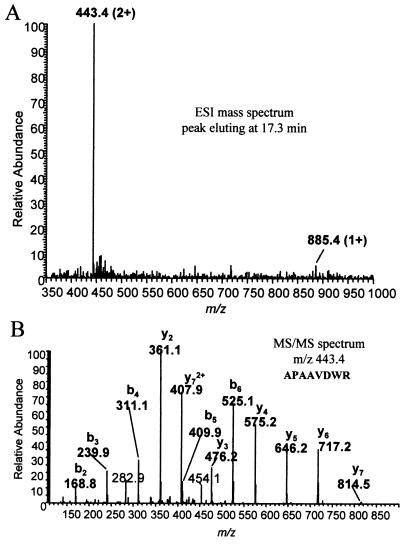
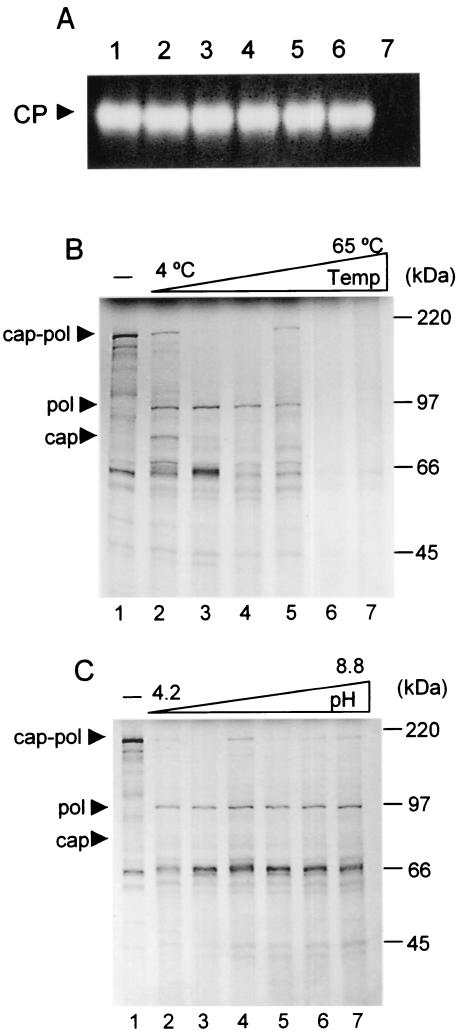
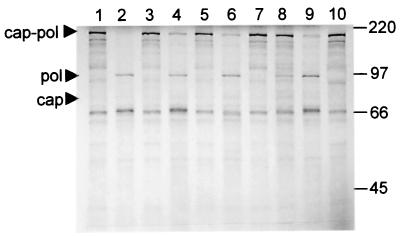
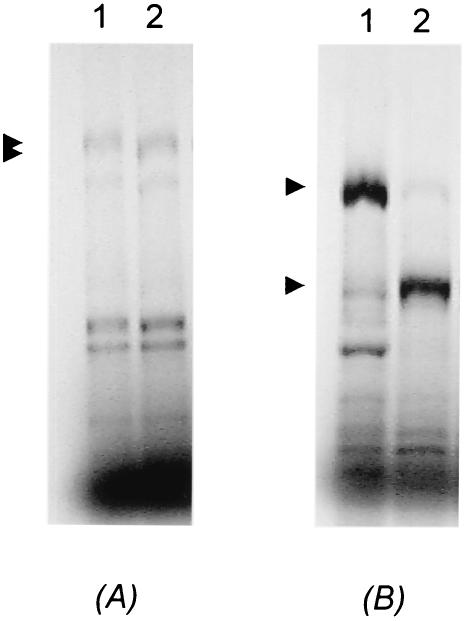
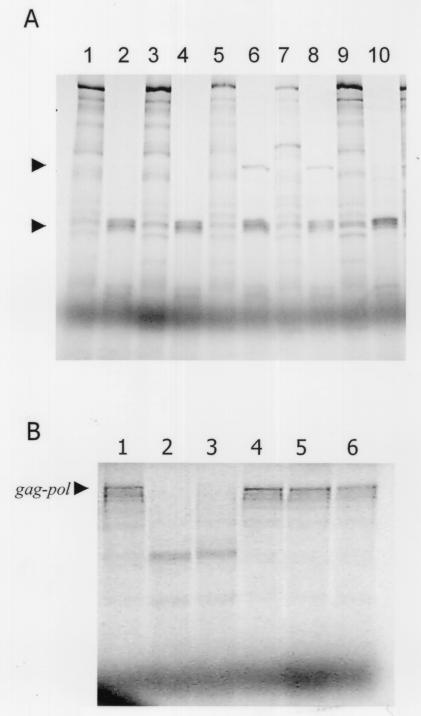
Leishmania RNA virus (LRV) is a double-stranded RNA virus that infects some strains of the protozoan parasite leishmania As with other totiviruses, LRV presumably expresses its polymerase by a ribosomal frameshift, resulting in a capsid-polymerase fusion protein. We have demonstrated previously that an LRV capsid-polymerase polyprotein is specifically cleaved by a Leishmania-encoded cysteine protease. This study reports the purification of this protease through a strategy involving anion-exchange chromatography and affinity chromatography. By using a Sepharose-immobilized lectin, concanavalin A, we isolated a fraction enriched with LRV polyprotein-specific protease activity. Analysis of the active fraction by sodium dodecyl sulfate-polyacrylamide gel electrophoreses and silver staining revealed a 50-kDa protein that, upon characterization by high-pressure liquid chromatography electrospray tandem mass spectrometry (electrospray ionization/MS/MS), was identified as a cysteine protease of trypanosomes. A partial amino acid sequence derived from the MS/MS data was compared with a protein database using BLAST software, revealing homology with several cysteine proteases of Leishmania and other trypanosomes. The protease exhibited remarkable temperature stability, while inhibitor studies characterized the protease as a trypsin-like cysteine protease-a novel finding for leishmania. To elucidate substrate preferences, a panel of deletion mutations and single-amino-acid mutations were engineered into a Gag-Pol fusion construct that was subsequently transcribed and translated in vitro and then used in cleavage assays. The data suggest that there are a number of cleavage sites located within a 153-amino-acid region spanning both the carboxy-terminal capsid region and the amino-terminal polymerase domain, with LRV capsid exhibiting the greatest susceptibility to proteolysis.
Figures

Similar articles
-
Specific in vitro cleavage of a Leishmania virus capsid-RNA-dependent RNA polymerase polyprotein by a host cysteine-like protease.J Virol. 1997 Dec;71(12):8983-90. doi: 10.1128/JVI.71.12.8983-8990.1997. J Virol. 1997. PMID: 9371554 Free PMC article.
-
Purification and biochemical characterization of a novel cysteine protease of Entamoeba histolytica.Eur J Biochem. 1999 Nov;266(1):170-80. doi: 10.1046/j.1432-1327.1999.00841.x. Eur J Biochem. 1999. PMID: 10542062
-
Single-site cleavage in the 5'-untranslated region of Leishmaniavirus RNA is mediated by the viral capsid protein.Proc Natl Acad Sci U S A. 1995 Sep 12;92(19):8994-8. doi: 10.1073/pnas.92.19.8994. Proc Natl Acad Sci U S A. 1995. PMID: 7568059 Free PMC article.
-
Overview of the Leishmaniavirus endoribonuclease and functions of other endoribonucleases affecting viral gene expression.J Exp Zool. 1998 Sep-Oct 1;282(1-2):254-60. J Exp Zool. 1998. PMID: 9723182 Review.
-
Roles of cysteine proteinases of trypanosomes and Leishmania in host-parasite interactions.Curr Opin Microbiol. 1998 Aug;1(4):455-60. doi: 10.1016/s1369-5274(98)80065-9. Curr Opin Microbiol. 1998. PMID: 10066510 Review.
Cited by
-
Relaxed cleavage specificity of an immunoglobulin A1 protease from Neisseria meningitidis.Infect Immun. 2007 Jun;75(6):2875-85. doi: 10.1128/IAI.01671-06. Epub 2007 Mar 12. Infect Immun. 2007. PMID: 17353288 Free PMC article.
-
Leishmania RNA virus: when the host pays the toll.Front Cell Infect Microbiol. 2012 Jul 12;2:99. doi: 10.3389/fcimb.2012.00099. eCollection 2012. Front Cell Infect Microbiol. 2012. PMID: 22919688 Free PMC article. Review.
-
Antiviral screening identifies adenosine analogs targeting the endogenous dsRNA Leishmania RNA virus 1 (LRV1) pathogenicity factor.Proc Natl Acad Sci U S A. 2017 Jan 31;114(5):E811-E819. doi: 10.1073/pnas.1619114114. Epub 2017 Jan 17. Proc Natl Acad Sci U S A. 2017. PMID: 28096399 Free PMC article.
References
-
- Barrett, A. J., and H. Kirschke. 1981. Cathepsin B, cathepsin H, and cathepsin L. Methods Enzymol. 80:535-561. - PubMed
-
- Bontempi, E., and J. J. Cazzulo. 1990. Digestion of human immunoglobulin G by the major cysteine proteinase (cruzipain) from Trypanosoma cruzi. FEMS Microbiol. Lett. 58:337-341. - PubMed
-
- Fujimura, T., J. C. Ribas, A. M. Makhov, and R. B. Wickner. 1992. Pol of gag-pol fusion protein required for encapsidation of viral RNA of yeast L-A virus. Nature 359:746-749. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
