

Does lowering glutamine synthetase activity in nodules modify nitrogen metabolism and growth of Lotus japonicus?
- PMID: 12970491
- PMCID: PMC196602
- DOI: 10.1104/pp.102.016766
Does lowering glutamine synthetase activity in nodules modify nitrogen metabolism and growth of Lotus japonicus?
Abstract
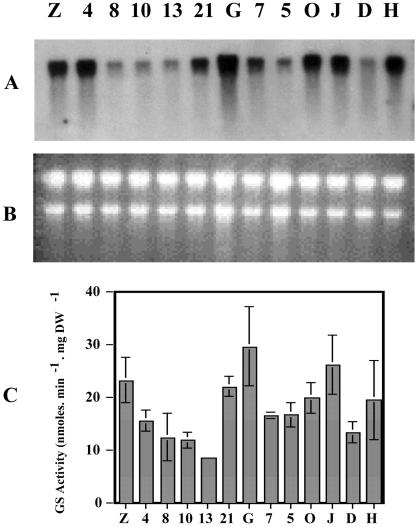
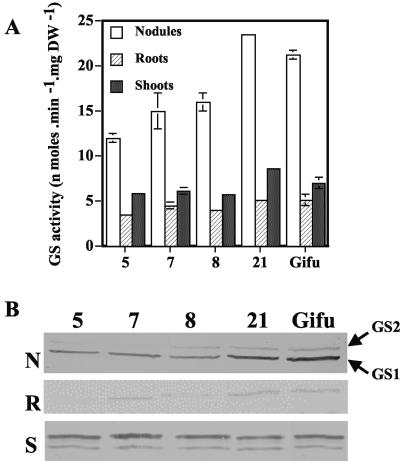
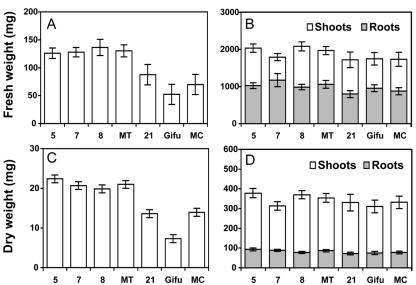
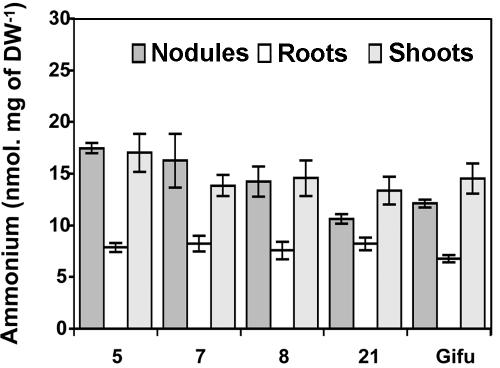
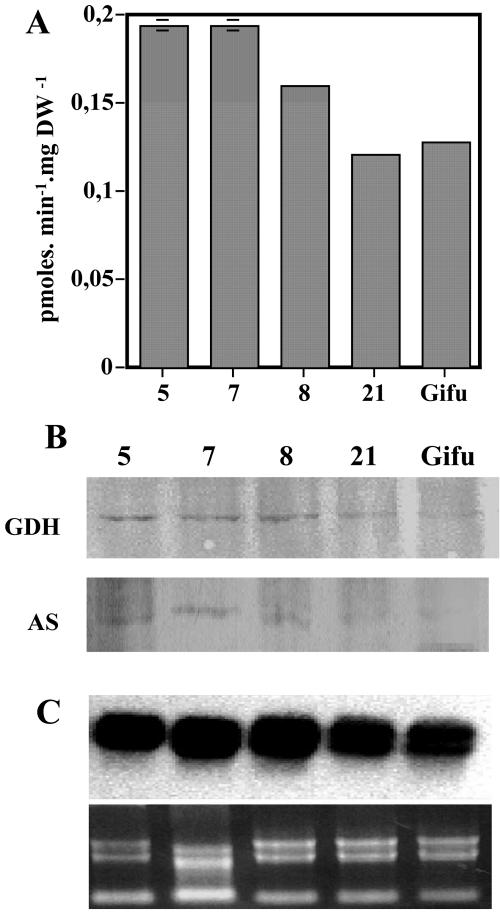
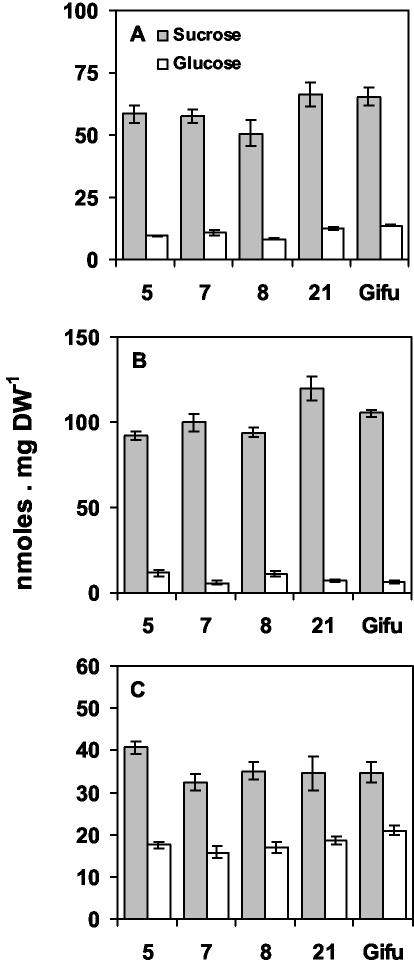
A cDNA encoding cytosolic glutamine synthetase (GS) from Lotus japonicus was fused in the antisense orientation relative to the nodule-specific LBC3 promoter of soybean (Glycine max) and introduced into L. japonicus via transformation with Agrobacterium tumefaciens. Among the 12 independent transformed lines into which the construct was introduced, some of them showed diminished levels of GS1 mRNA and lower levels of GS activity. Three of these lines were selected and their T(1) progeny was further analyzed both for plant biomass production and carbon and nitrogen (N) metabolites content under symbiotic N-fixing conditions. Analysis of these plants revealed an increase in fresh weight in nodules, roots and shoots. The reduction in GS activity was found to correlate with an increase in amino acid content of the nodules, which was primarily due to an increase in asparagine content. Thus, this study supports the hypothesis that when GS becomes limiting, other enzymes (e.g. asparagine synthetase) that have the capacity to assimilate ammonium may be important in controlling the flux of reduced N in temperate legumes such as L. japonicus. Whether these alternative metabolic pathways are important in the control of plant biomass production still remains to be fully elucidated.
Figures
References
-
- Becker D, Kemper E, Schell J, Masterson R (1992a) New plant binary vectors with selectable markers located proximal to the left T-DNA border. Plant Mol Biol 20: 1195-1197 - PubMed
-
- Becker TW, Caboche M, Carrayol E, Hirel B (1992b) Nucleotide sequence of a tobacco cDNA encoding plastidic glutamine synthetase and light-inducibility, organ specificity and diurnal rhythmicity in the expression of the corresponding genes of tobacco and tomato. Plant Mol Biol 19: 367-379 - PubMed
-
- Beevers L (1976) Nitrogen Metabolism in Plants. EJW Barrington, AJ Willis, eds, E Arnold, London, UK pp 1-333
-
- Bergmeyer HU (1974). Methods of Enzymatic Analysis, Vol. 3. Academic Press, New York, pp 1176-1179
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
