

The association between integrin-associated protein and SHPS-1 regulates insulin-like growth factor-I receptor signaling in vascular smooth muscle cells
- PMID: 12972543
- PMCID: PMC196546
- DOI: 10.1091/mbc.e03-04-0239
The association between integrin-associated protein and SHPS-1 regulates insulin-like growth factor-I receptor signaling in vascular smooth muscle cells
Abstract
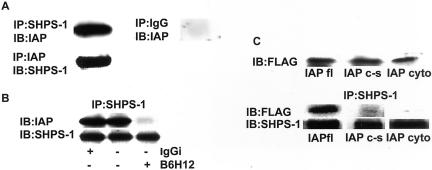
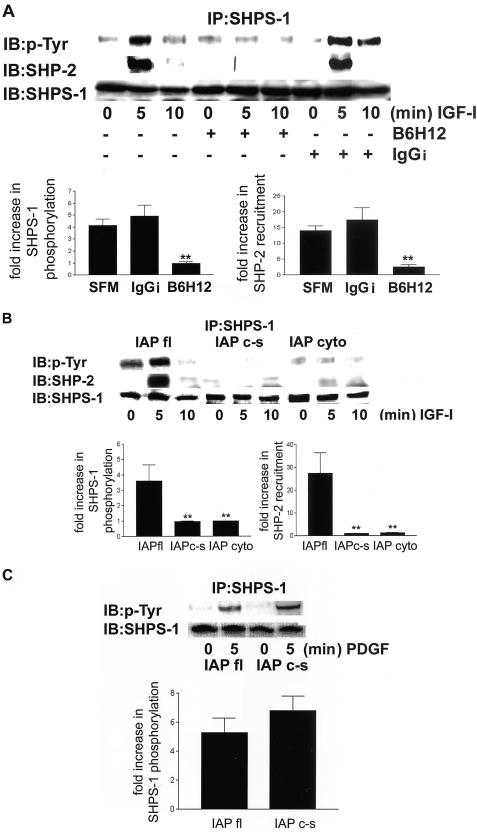
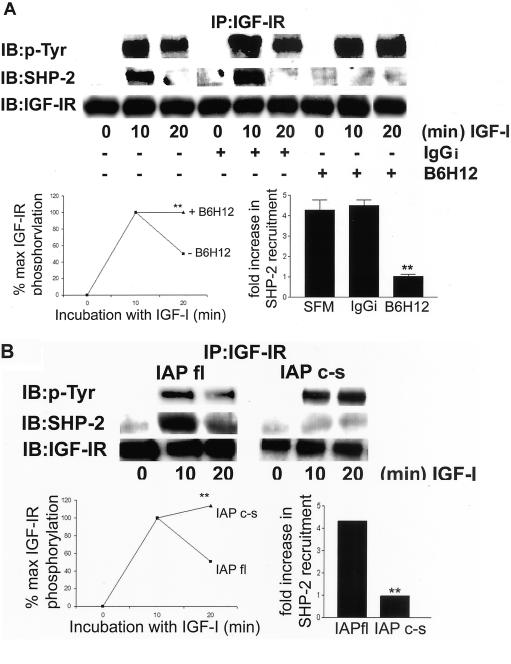
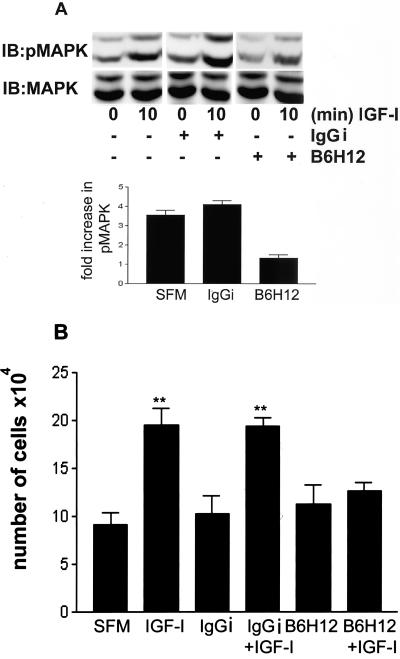
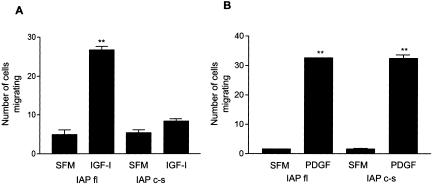
Growth factor signaling is usually analyzed in isolation without considering the effect of ligand occupancy of transmembrane proteins other than the growth factor receptors themselves. In smooth muscle cells, the transmembrane protein Src homology 2 domain containing protein tyrosine phosphatase substrate-1 (SHPS-1) has been shown to be an important regulator of insulin-like growth factor-I (IGF-I) signaling. SHPS-1 is phosphorylated in response to IGF-I, leading to recruitment of Src homology 2 domain tyrosine phosphatase (SHP-2). Subsequently, SHP-2 is transferred to IGF-I receptor and regulates the duration of IGF-I receptor phosphorylation. Whether ligand occupancy of SHPS-1 influences SHPS-1 phosphorylation or SHP-2 recruitment, thereby altering growth factor signaling, is unknown. Previous studies have shown that integrin associated protein (IAP) associates with SHPS-1. We undertook these studies to determine whether this interaction controlled SHPS-1 phosphorylation and/or SHP-2 recruitment and thereby regulated IGF-I signaling. Disruption of IAP-SHPS-1 binding, by using an IAP monoclonal antibody or cells expressing mutant forms of IAP that did not bind to SHPS-1, inhibited IGF-I-stimulated SHPS-1 phosphorylation and SHP-2 recruitment. This was associated with a lack of SHP-2 transfer to IGF-I receptor and sustained receptor phosphorylation. This resulted in an inability of IGF-I to stimulate sustained mitogen-activated protein kinase activation, cell proliferation, and cell migration. The effect was specific for IGF-I because disruption of the IAP-SHPS-1 interaction had no effect on platelet-derived growth factor-stimulated SHPS-1 phosphorylation or cell migration. In summary, our results show that 1) ligand occupancy of SHPS-1 is a key determinant of its ability to be phosphorylated after IGF-I stimulation, and 2) the interaction between IAP and SHPS-1 is an important regulator of IGF-I signaling because disruption of the results in impaired SHP-2 recruitment and subsequent inhibition of IGF-I-stimulated cell proliferation and migration.
Figures
References
-
- Andre, F., Rigot, V., Thimonier, J., Montixi, C., Parat, F., Pommier, G., Marvaldi, J., and Luis, J. (1999). Integrins and E-cadherin cooperate with IGF-I to induce migration of epithelial colonic cells. Int. J. Cancer 83, 497–505. - PubMed
-
- Babic, I., Schallhorn, A., Lindberg, F.P., and Jirik, F.R. (2000). SHPS-1 induces aggregation of Ba/F3 pro-B cells via an interaction with CD47. J. Immunol. 164, 3652–3658. - PubMed
-
- Fujioka, Y., Matozaki, T., Noguchi, T., Iwamatsu, A., Yamao, T., Takahashi, N., Tsuda, M., Takada, T., and Kasuga, M. (1996). A novel membrane glycoprotein, SHPS-1, that binds the SH2-domain-containing protein tyrosine phosphatase SHP-2 in response to mitogens and cell adhesion. Mol. Cell. Biol. 16, 6887–6899. - PMC - PubMed
-
- Gockerman, A., Prevette, T., Jones, J.I., and Clemmons, D.R. (1995). Insulin-like growth factor (IGF)-binding proteins inhibit the smooth muscle cell migration responses to IGF-I and IGF-II. Endocrinology 136, 4168–4173. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
