

RAM: a conserved signaling network that regulates Ace2p transcriptional activity and polarized morphogenesis
- PMID: 12972564
- PMCID: PMC196567
- DOI: 10.1091/mbc.e03-01-0018
RAM: a conserved signaling network that regulates Ace2p transcriptional activity and polarized morphogenesis
Abstract
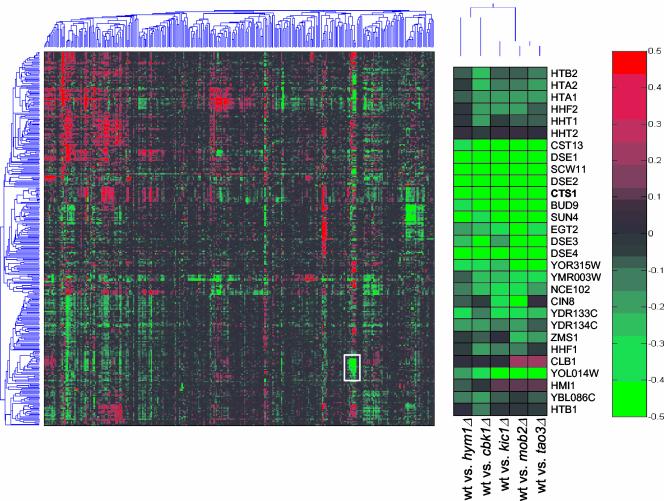
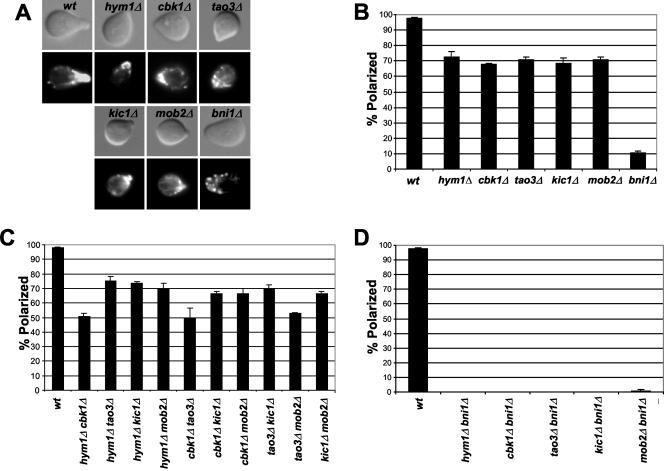
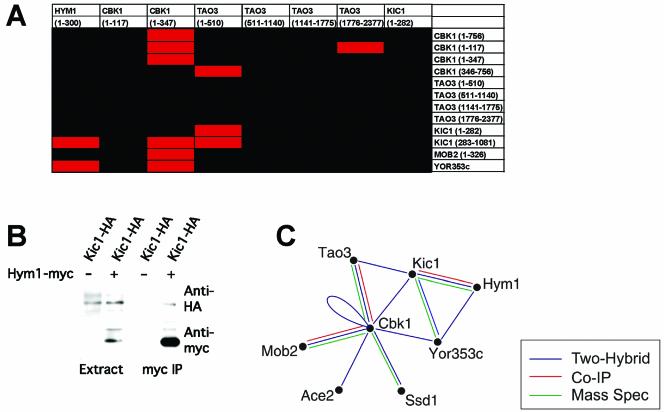
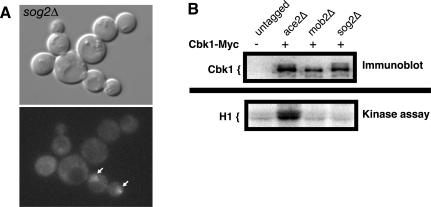
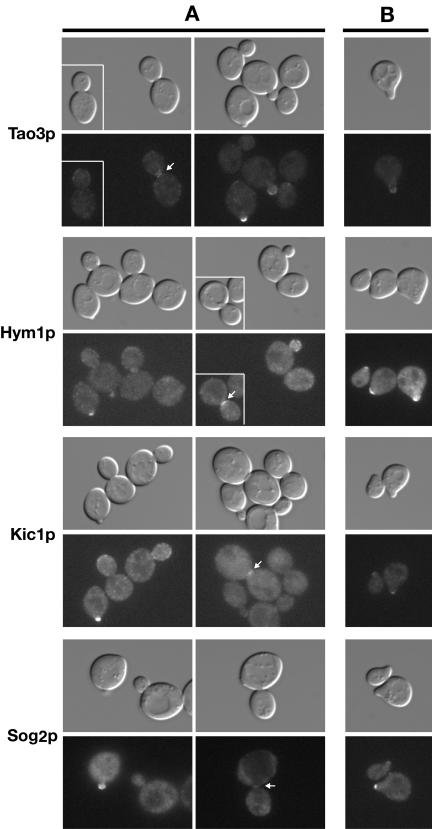
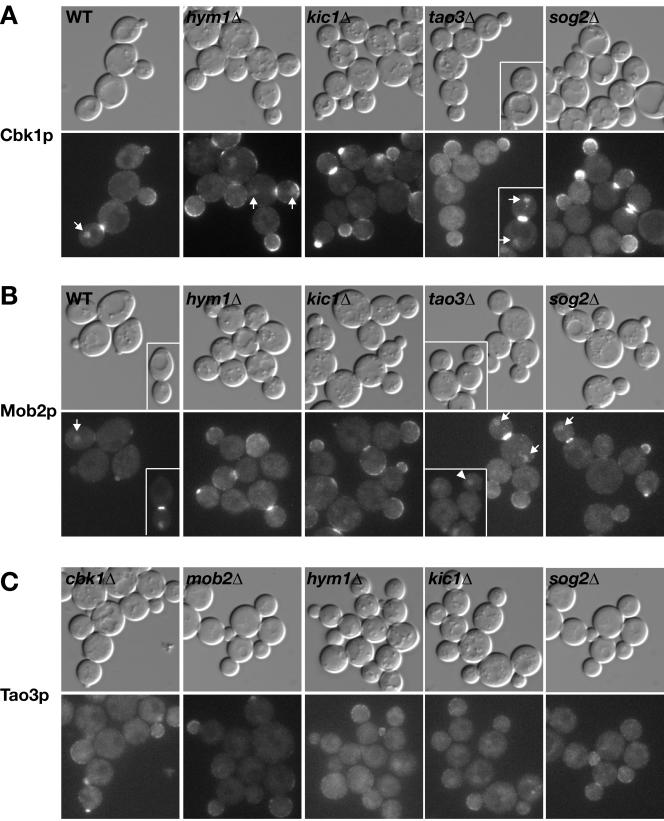
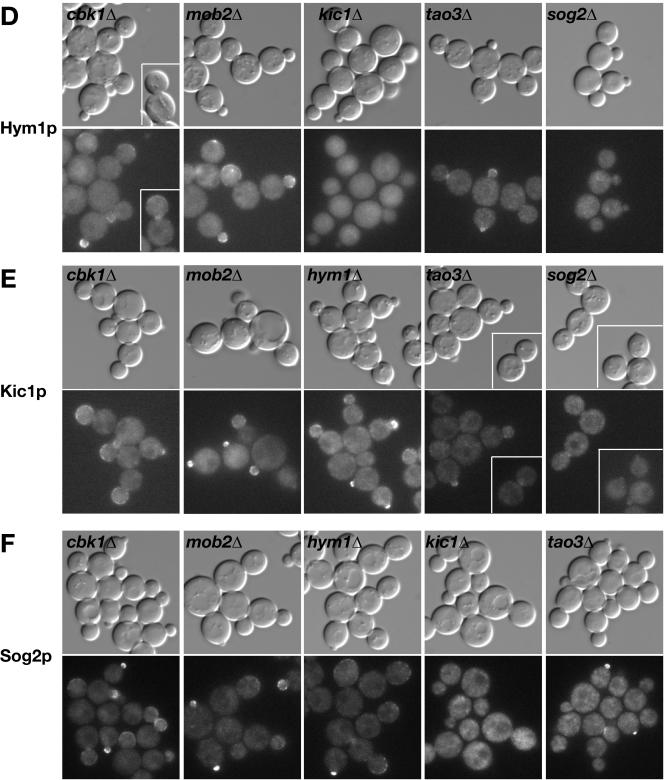
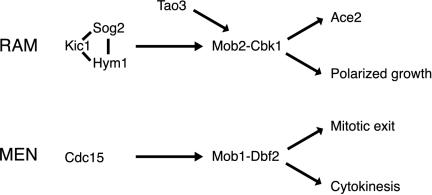
In Saccharomyces cerevisiae, polarized morphogenesis is critical for bud site selection, bud development, and cell separation. The latter is mediated by Ace2p transcription factor, which controls the daughter cell-specific expression of cell separation genes. Recently, a set of proteins that include Cbk1p kinase, its binding partner Mob2p, Tao3p (Pag1p), and Hym1p were shown to regulate both Ace2p activity and cellular morphogenesis. These proteins seem to form a signaling network, which we designate RAM for regulation of Ace2p activity and cellular morphogenesis. To find additional RAM components, we conducted genetic screens for bilateral mating and cell separation mutants and identified alleles of the PAK-related kinase Kic1p in addition to Cbk1p, Mob2p, Tao3p, and Hym1p. Deletion of each RAM gene resulted in a loss of Ace2p function and caused cell polarity defects that were distinct from formin or polarisome mutants. Two-hybrid and coimmunoprecipitation experiments reveal a complex network of interactions among the RAM proteins, including Cbk1p-Cbk1p, Cbk1p-Kic1p, Kic1p-Tao3p, and Kic1p-Hym1p interactions, in addition to the previously documented Cbk1p-Mob2p and Cbk1p-Tao3p interactions. We also identified a novel leucine-rich repeat-containing protein Sog2p that interacts with Hym1p and Kic1p. Cells lacking Sog2p exhibited the characteristic cell separation and cell morphology defects associated with perturbation in RAM signaling. Each RAM protein localized to cortical sites of growth during both budding and mating pheromone response. Hym1p was Kic1p- and Sog2p-dependent and Sog2p and Kic1p were interdependent for localization, indicating a close functional relationship between these proteins. Only Mob2p and Cbk1p were detectable in the daughter cell nucleus at the end of mitosis. The nuclear localization and kinase activity of the Mob2p-Cbk1p complex were dependent on all other RAM proteins, suggesting that Mob2p-Cbk1p functions late in the RAM network. Our data suggest that the functional architecture of RAM signaling is similar to the S. cerevisiae mitotic exit network and Schizosaccharomyces pombe septation initiation network and is likely conserved among eukaryotes.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
