

Pancreatic-duodenal homeobox 1 regulates expression of liver receptor homolog 1 during pancreas development
- PMID: 12972592
- PMCID: PMC193920
- DOI: 10.1128/MCB.23.19.6713-6724.2003
Pancreatic-duodenal homeobox 1 regulates expression of liver receptor homolog 1 during pancreas development
Abstract
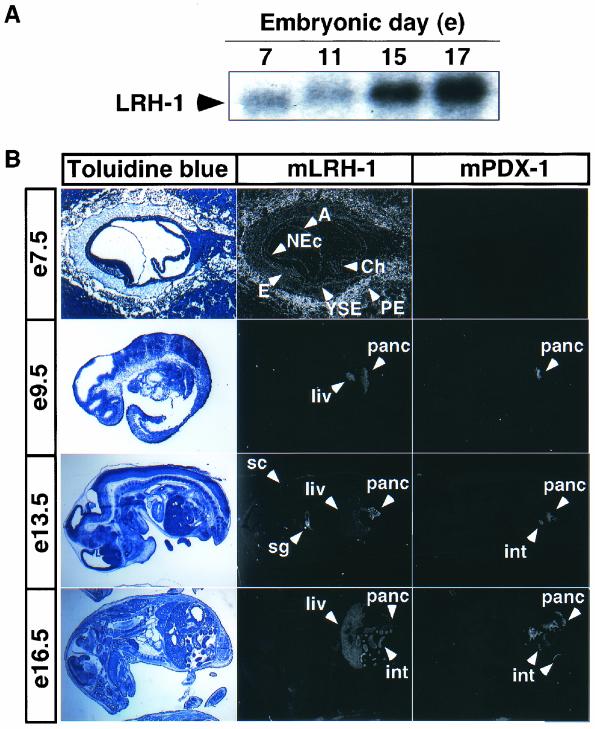
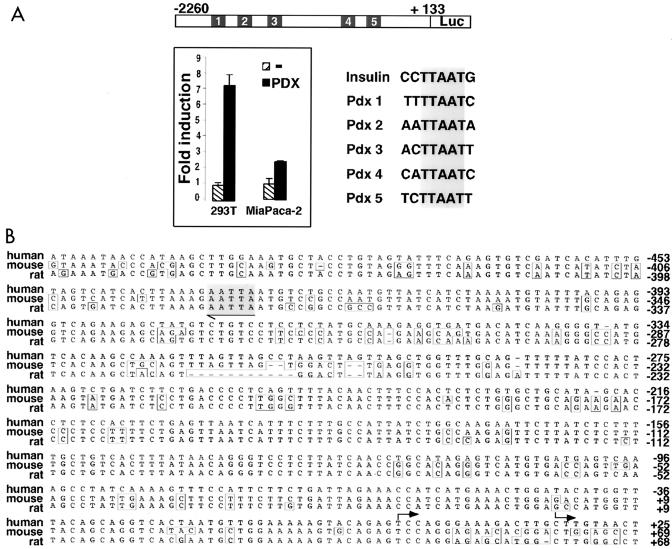
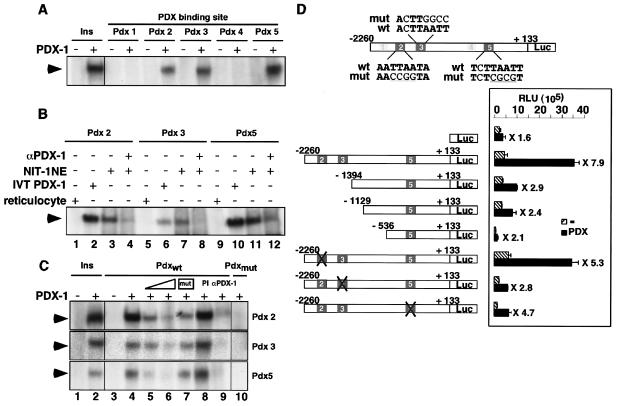
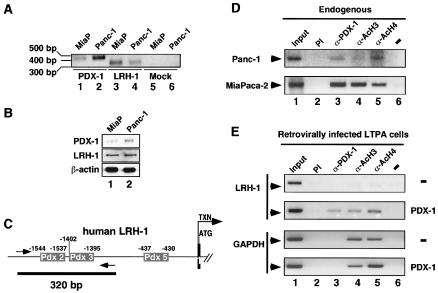
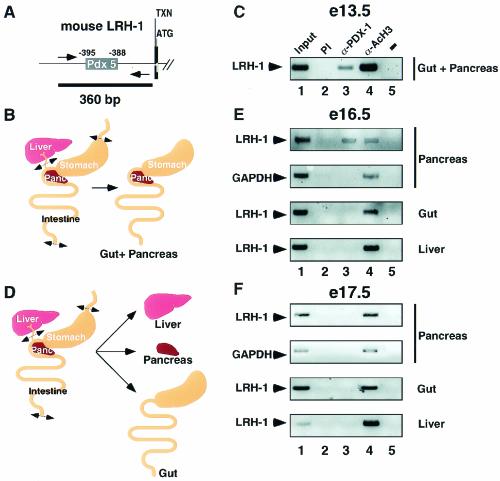
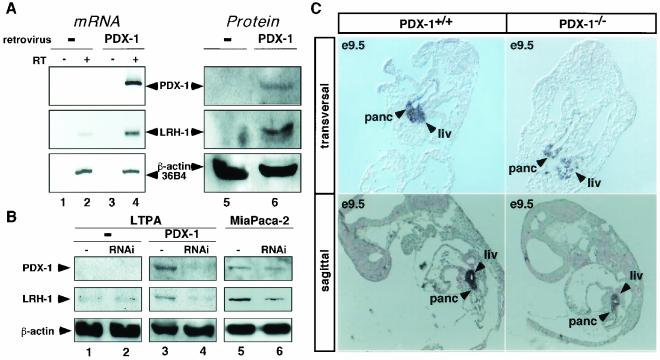
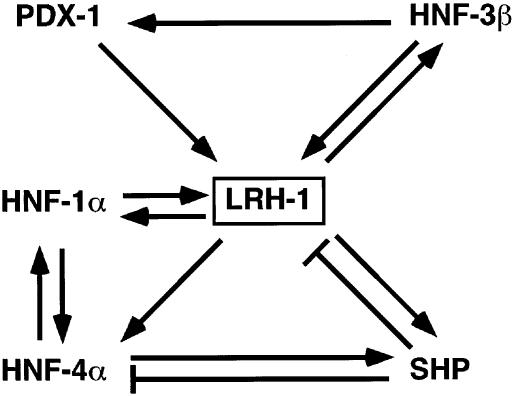
Liver receptor homolog 1 (LRH-1) and pancreatic-duodenal homeobox 1 (PDX-1) are coexpressed in the pancreas during mouse embryonic development. Analysis of the regulatory region of the human LRH-1 gene demonstrated the presence of three functional binding sites for PDX-1. Electrophoretic mobility shift assays and chromatin immunoprecipitation analysis showed that PDX-1 bound to the LRH-1 promoter, both in cultured cells in vitro and during pancreatic development in vivo. Retroviral expression of PDX-1 in pancreatic cells induced the transcription of LRH-1, whereas reduced PDX-1 levels by RNA interference attenuated its expression. Consistent with direct regulation of LRH-1 expression by PDX-1, PDX-1(-/-) mice expressed smaller amounts of LRH-1 mRNA in the embryonic pancreas. Taken together, our data indicate that PDX-1 controls LRH-1 expression and identify LRH-1 as a novel downstream target in the PDX-1 regulatory cascade governing pancreatic development, differentiation, and function.
Figures
References
-
- Ahlgren, U., J. Jonsson, and H. Edlund. 1996. The morphogenesis of the pancreatic mesenchyme is uncoupled from that of the pancreatic epithelium in IPF1/PDX1-deficient mice. Development 122:1409-1416. - PubMed
-
- Ang, S. L., and J. Rossant. 1994. HNF-3 beta is essential for node and notochord formation in mouse development. Cell 78:561-574. - PubMed
-
- Becker-André, M., E. André, and J. F. DeLamarter 1993. Identification of nuclear receptor mRNAs by RT-PCR amplification of conserved zinc-finger motif sequences. Biochem. Biophys. Res. Commun. 194:1371-1379. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials