

A phosphomimetic mutation at Ser-138 renders iron regulatory protein 1 sensitive to iron-dependent degradation
- PMID: 12972614
- PMCID: PMC193948
- DOI: 10.1128/MCB.23.19.6973-6981.2003
A phosphomimetic mutation at Ser-138 renders iron regulatory protein 1 sensitive to iron-dependent degradation
Abstract
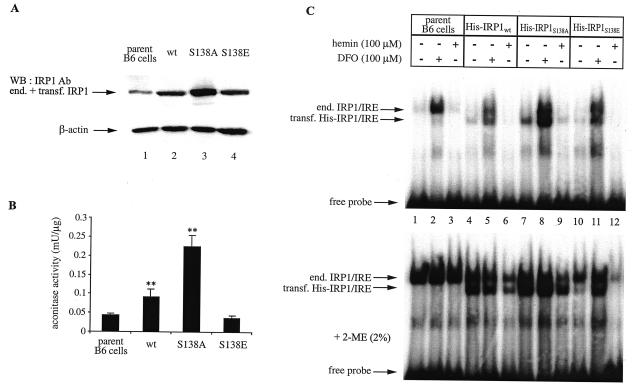
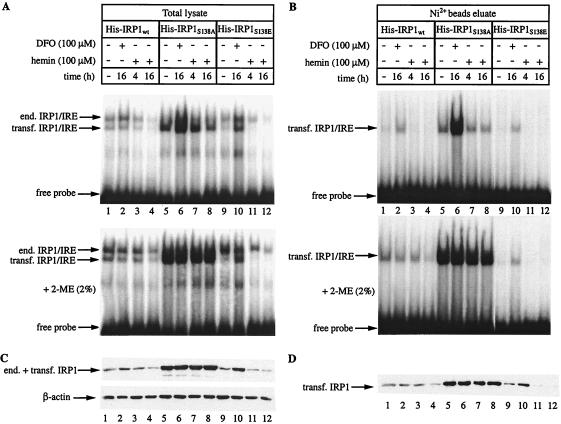
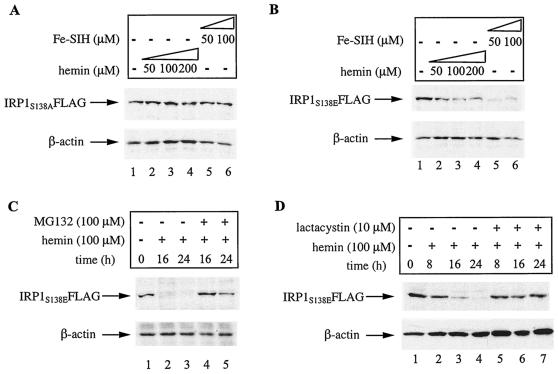
Iron regulatory protein 1 (IRP1) binds to mRNA iron-responsive elements (IREs) and thereby controls the expression of IRE-containing mRNAs. In iron-replete cells, assembly of a cubane [4Fe-4S] cluster inhibits IRE-binding activity and converts IRP1 to a cytosolic aconitase. Earlier experiments with Saccharomyces cerevisiae suggested that phosphomimetic mutations of Ser-138 negatively affect the stability of the cluster (N. M. Brown, S. A. Anderson, D. W. Steffen, T. B. Carpenter, M. C. Kennedy, W. E. Walden, and R. S. Eisenstein, Proc. Natl. Acad. Sci. USA 95:15235-15240, 1998). Along these lines, we show here that a highly purified preparation of recombinant human IRP1 bearing a phosphomimetic S138E substitution (IRP1(S138E)) lacks aconitase activity, which is a hallmark of [4Fe-4S] cluster integrity. Similarly, IRP1(S138E) expressed in mammalian cells fails to function as aconitase. Furthermore, we demonstrate that the impairment of [4Fe-4S] cluster assembly in mammalian cells sensitizes IRP1(S138E) to iron-dependent degradation. This effect can be completely blocked by the iron chelator desferrioxamine or by the proteasome inhibitors MG132 and lactacystin. As expected, the stability of wild-type or phosphorylation-deficient IRP1(S138A) is not affected by iron manipulations. Ser-138 and flanking sequences appear to be highly conserved in the IRP1s of vertebrates, whereas insect IRP1 orthologues and nonvertebrate IRP1-like molecules contain an S138A substitution. Our data suggest that phosphorylation of Ser-138 may provide a basis for an additional mechanism for the control of vertebrate IRP1 activity at the level of protein stability.
Figures




References
-
- Beinert, H., R. H. Holm, and E. Münck. 1997. Iron-sulfur clusters: nature's modular, multipurpose structures. Science 277:653-659. - PubMed
-
- Brazzolotto, X., J. Gaillard, K. Pantopoulos, M. W. Hentze, and J.-M. Moulis. 1999. Human cytoplasmic aconitase (iron regulatory protein 1) is converted into its [3Fe-4S] form by hydrogen peroxide in vitro but is not activated for iron-responsive element binding. J. Biol. Chem. 274:21625-21630. - PubMed
-
- Brown, N. M., S. A. Anderson, D. W. Steffen, T. B. Carpenter, M. C. Kennedy, W. E. Walden, and R. S. Eisenstein. 1998. Novel role of phosphorylation in Fe-S cluster stability revealed by phosphomimetic mutations at Ser-138 of iron regulatory protein 1. Proc. Natl. Acad. Sci. USA 95:15235-15240. - PMC - PubMed
-
- Brown, N. M., M. C. Kennedy, W. E. Antholine, R. S. Eisenstein, and W. E. Walden. 2002. Detection of a [3Fe-4S] cluster intermediate of cytosolic aconitase in yeast expressing iron regulatory protein 1: insights into the mechanism of Fe-S cluster cycling. J. Biol. Chem. 277:7246-7254. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials