

Analysis of the wild-type and mutant genes encoding the enzyme kynurenine monooxygenase of the yellow fever mosquito, Aedes aegypti
- PMID: 12974953
- PMCID: PMC2629591
- DOI: 10.1046/j.1365-2583.2003.00433.x
Analysis of the wild-type and mutant genes encoding the enzyme kynurenine monooxygenase of the yellow fever mosquito, Aedes aegypti
Abstract
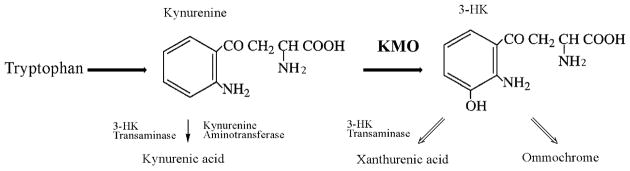
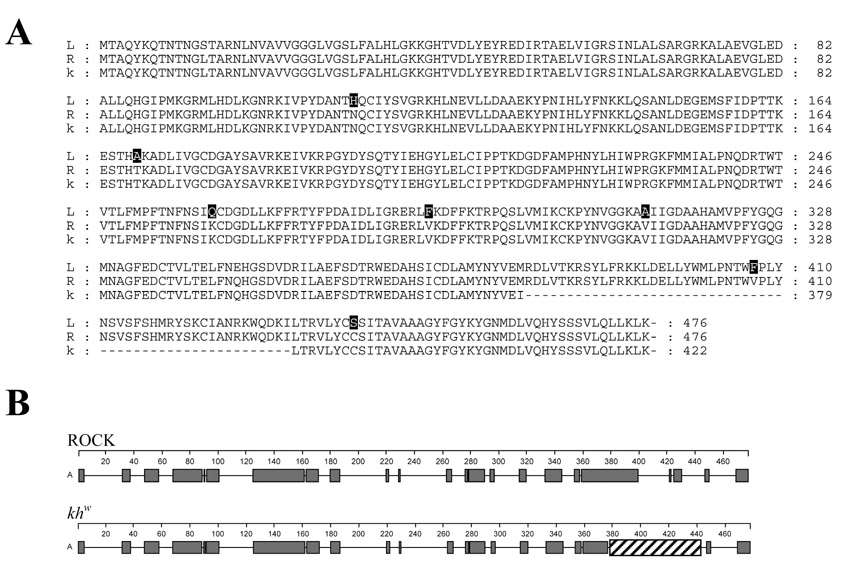
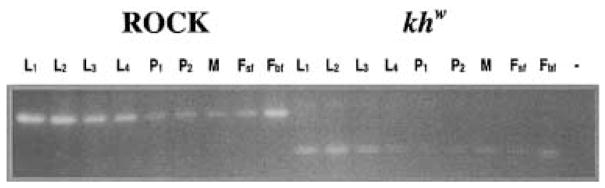
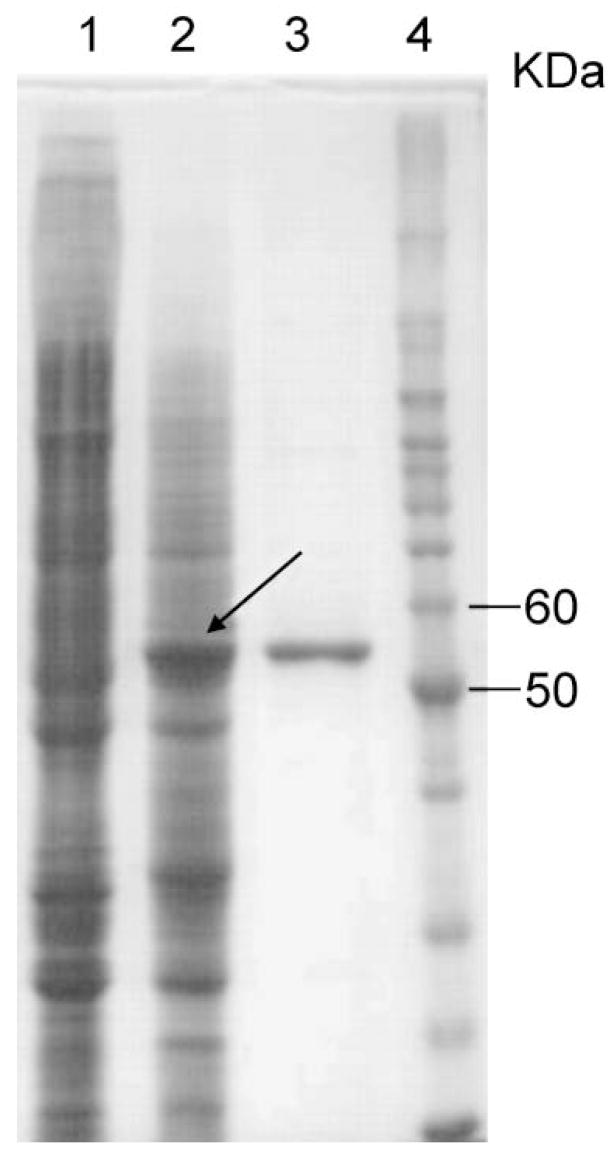
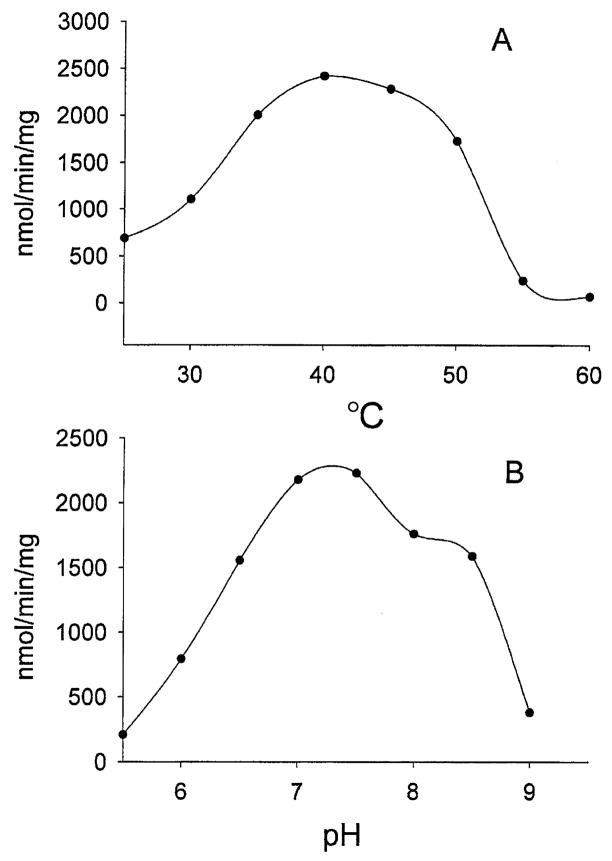
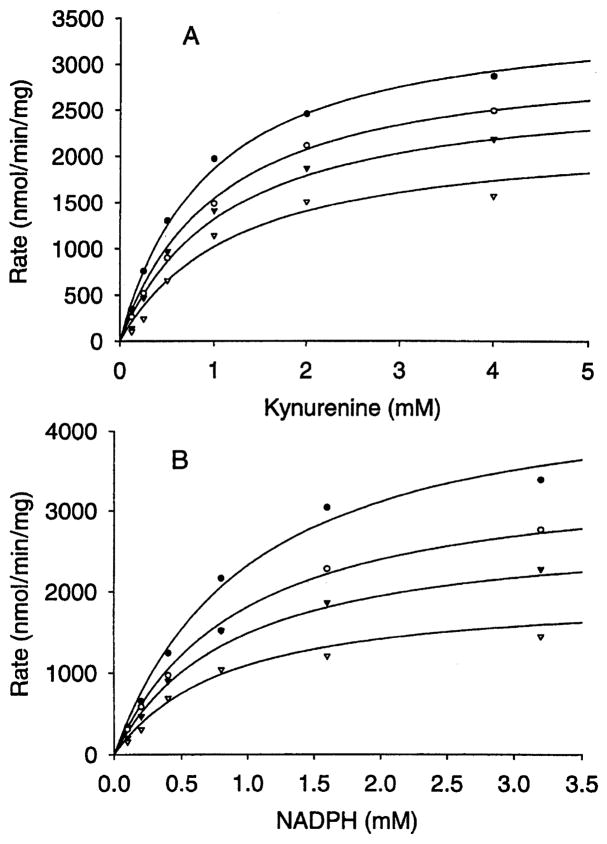
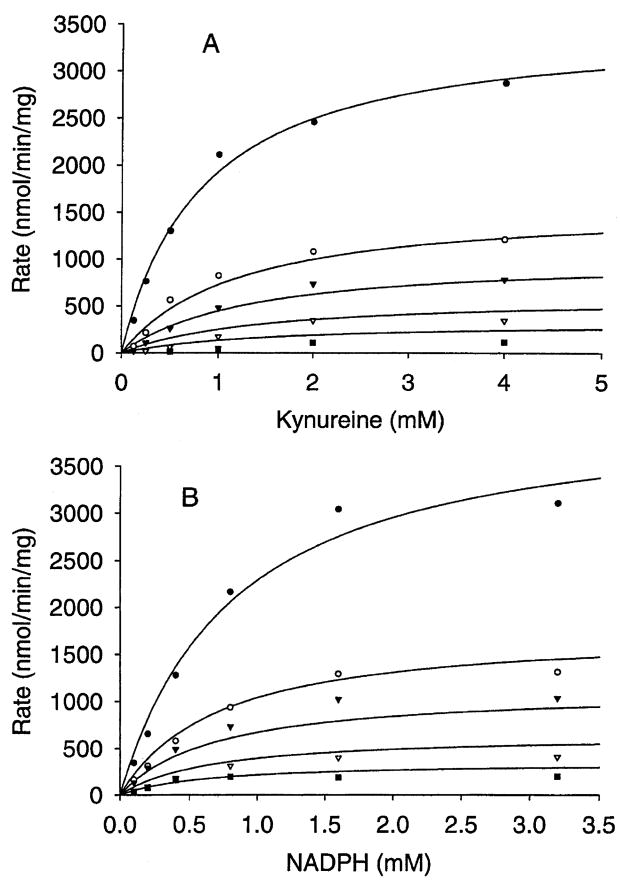
Kynurenine 3-monooxygenase (KMO) catalyses the hydroxylation of kynurenine to 3-hydroxykynurenine. KMO has a key role in tryptophan catabolism and synthesis of ommochrome pigments in mosquitoes. The gene encoding this enzyme in the yellow fever mosquito, Aedes aegypti, is called kynurenine hydroxylase (kh) and a mutant allele that produces white eyes has been designated khw. A number of cDNA clones representative of wild-type and mutant genes were isolated. Sequence analyses of the wild-type and mutant cDNAs revealed a deletion of 162 nucleotides in the mutant gene near the 3'-end of the deduced coding region. RT-PCR analyses confirm the transcription of a truncated mRNA in the mutant strain. The in-frame deletion results in a loss of 54 amino acids, which disrupts a major alpha-helix and which probably accounts for the loss of activity of the enzyme. Recombinant Ae. aegypti KMO showed high substrate specificity for kynurenine with optimum activity at 40 degrees C and pH = 7.5. Kinetic parameters and inhibition of KMO activity by Cl- and pyridoxal-5-phosphate were determined.
Figures

References
-
- Akaboshi E. Kynurenine hydroxylase in Musca domestica L. Comp Biochem Physiol B. 1979;62:549–555. - PubMed
-
- Arai M, Billker O, Morris HR, Panico M, Delcroix M, Dixon D, Ley SV, Sinden RE. Both mosquito-derived xanthurenic acid and a host blood-derived factor regulate gametogenesis of Plasmodium in the midgut of the mosquito. Mol Biochem Parasitol. 2001;116:17–24. - PubMed
-
- Atkinson PW, Pinkerton AC, O’Brochta DA. Genetic transformation systems in insects. Annu Rev Entomol. 2001;46:317–346. - PubMed
-
- Bhalla SC. White eye, a new sex-linked mutant of Aedes aegypti. Mosquito News. 1968;28:380–385.
-
- Billker O, Lindo V, Panico M, Etienne AE, Paxton T, Dell A, Rogers M, Sinden RE, Morris HR. Identification of xanthurenic acid as the putative inducer of malaria development in the mosquito. Nature. 1998;392:289–292. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
