

Ku interacts with telomerase RNA to promote telomere addition at native and broken chromosome ends
- PMID: 12975323
- PMCID: PMC218076
- DOI: 10.1101/gad.1125903
Ku interacts with telomerase RNA to promote telomere addition at native and broken chromosome ends
Abstract
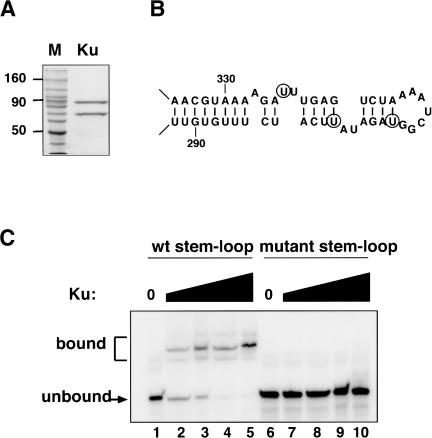
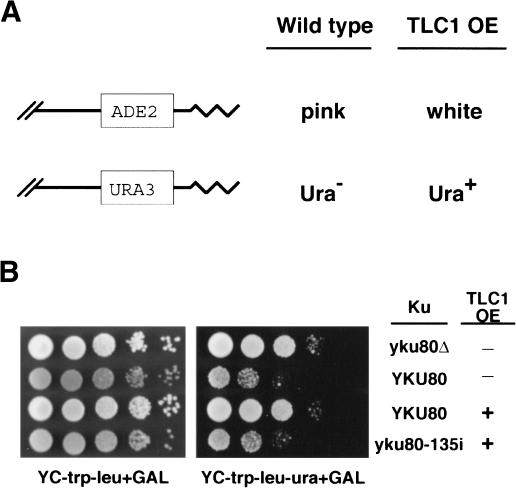
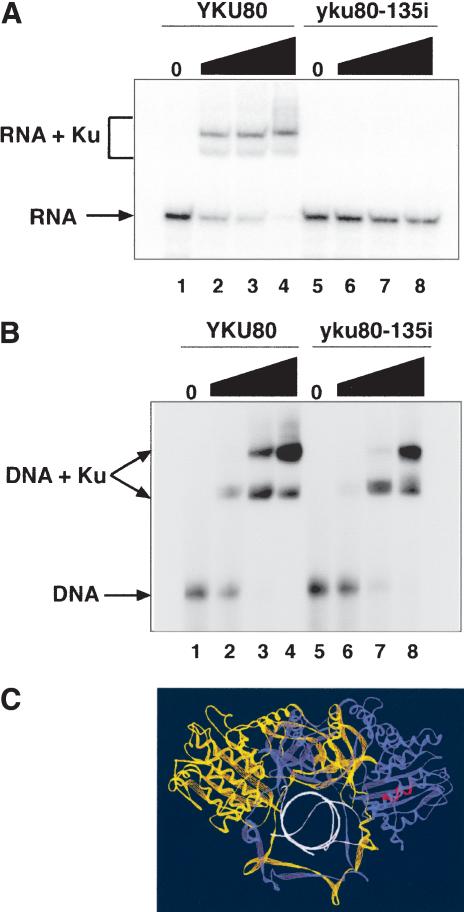
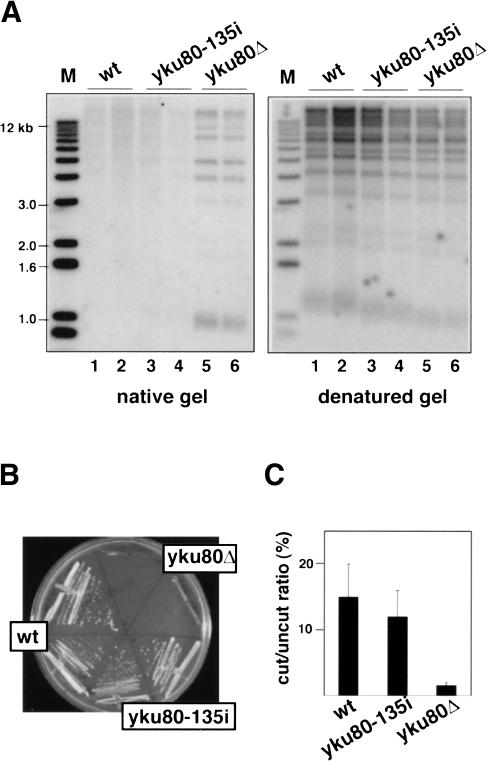
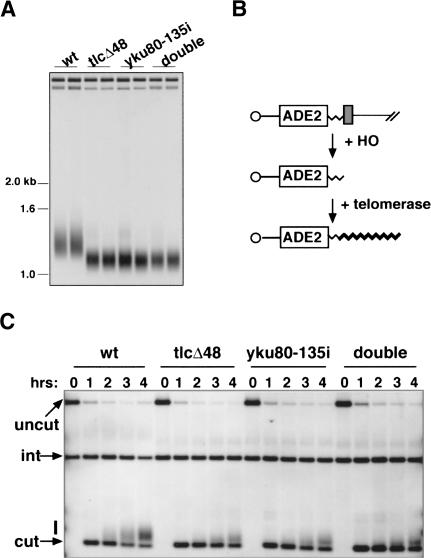
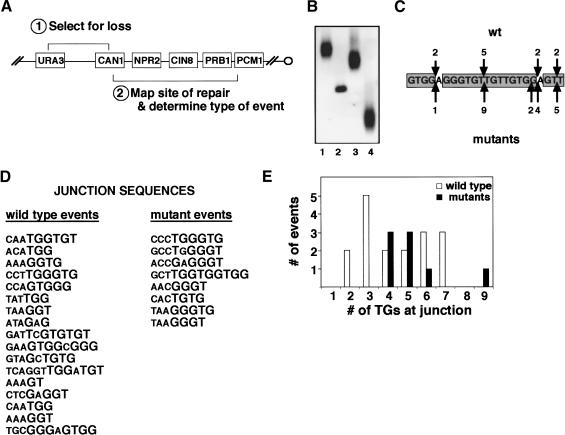
Ku is a conserved DNA end-binding protein that plays various roles at different kinds of DNA ends. At telomeres, Ku is part of the structure that protects the chromosome end, whereas at broken DNA ends, Ku promotes DNA repair as part of the nonhomologous end-joining (NHEJ) pathway. Here, we present evidence of a new role for Ku that impacts both telomere-length maintenance and DNA repair in Saccharomyces cerevisiae. We show that Ku binds TLC1, the RNA component of telomerase. We also describe a novel separation-of-function allele of Ku that is specifically defective in TLC1 binding. In this mutant, telomeres are short and the kinetics of telomere addition are slow, but other Ku-dependent activities, such as chromosome end protection and NHEJ, are unaffected. At low frequency, yeast will use telomerase to heal DNA damage by capping the broken chromosome with telomeric DNA sequences. We show that when Ku's ability to bind TLC1 is disrupted, DNA repair via telomere healing is reduced 10- to 100-fold, and the spectrum of sequences that can acquire a telomere changes. Thus, the interaction between Ku and TLC1 RNA enables telomerase to act at both broken and normal chromosome ends.
Figures
Comment in
-
Which end: dissecting Ku's function at telomeres and double-strand breaks.Genes Dev. 2003 Oct 1;17(19):2347-50. doi: 10.1101/gad.1146603. Genes Dev. 2003. PMID: 14522942 Review. No abstract available.
References
-
- Anderson E.M., Halsey, W.A., and Wuttke, D.S. 2003. Site-directed mutagenesis reveals the thermodynamic requirements for single-stranded DNA recognition by the telomere-binding protein Cdc13. Biochemistry 42: 3751-3758. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous