

The budding yeast silencing protein Sir1 is a functional component of centromeric chromatin
- PMID: 12975325
- PMCID: PMC218072
- DOI: 10.1101/gad.1131103
The budding yeast silencing protein Sir1 is a functional component of centromeric chromatin
Abstract
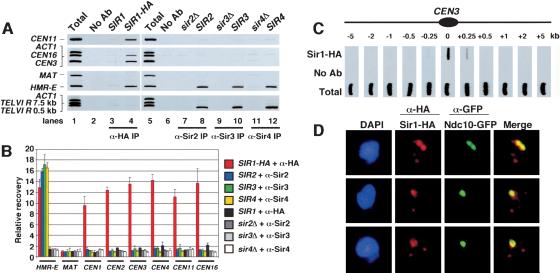
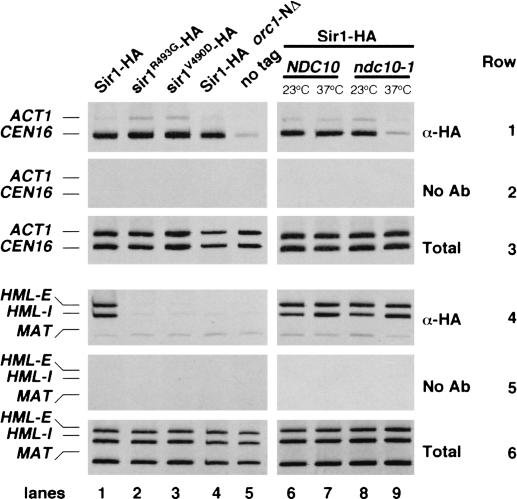
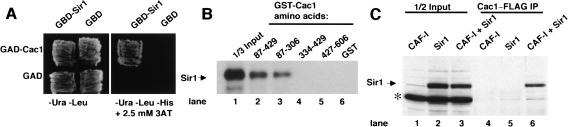
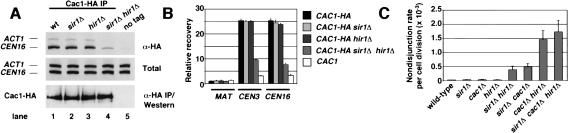
In fission yeast and multicellular organisms, centromere-proximal regions of chromosomes are heterochromatic, containing proteins that silence gene expression. In contrast, the relationship between heterochromatin proteins and kinetochore function in the budding yeast Saccharomyces cerevisiae remains largely unexplored. Here we report that the yeast heterochromatin protein Sir1 is a component of centromeric chromatin and contributes to mitotic chromosome stability. Sir1 recruitment to centromeres occurred through a novel mechanism independent of its interaction with the origin recognition complex (ORC). Sir1 function at centromeres was distinct from its role in forming heterochromatin, because the Sir2-4 proteins were not associated with centromeric regions. Sir1 bound to Cac1, a subunit of chromatin assembly factor I (CAF-I), and helped to retain Cac1 at centromeric loci. These studies reveal that although budding yeast and mammalian cells use fundamentally different mechanisms of forming heterochromatin, they both use silencing proteins to attract the histone deposition factor CAF-I to centromeric chromatin.
Figures
References
-
- Bernard P., Maura, J.F., Partridge, J.F., Genier, S., Javerzat, J.P., and Allshire, R.C. 2001. Requirement of heterochromatin for cohesion at centromeres. Science 294: 2539-2542. - PubMed
-
- Biggins S. and Walczak, C.E. 2003. Captivating capture: How microtubules attach to kinetochores. Curr. Biol. 13: R449-R460. - PubMed
-
- Bryk M., Briggs, S.D., Strahl, B.D., Curcio, M.J., Allis, C.D., and Winston, F. 2002. Evidence that Set1, a factor required for methylation of histone H3, regulates rDNA silencing in S. cerevisiae by a Sir2-independent mechanism. Curr. Biol. 12: 165-170. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases