

Calsequestrin determines the functional size and stability of cardiac intracellular calcium stores: Mechanism for hereditary arrhythmia
- PMID: 13130076
- PMCID: PMC208831
- DOI: 10.1073/pnas.1932318100
Calsequestrin determines the functional size and stability of cardiac intracellular calcium stores: Mechanism for hereditary arrhythmia
Abstract
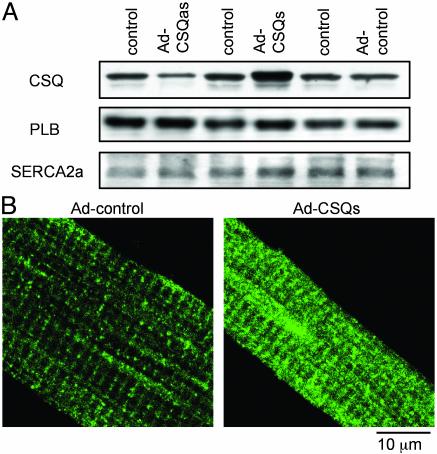
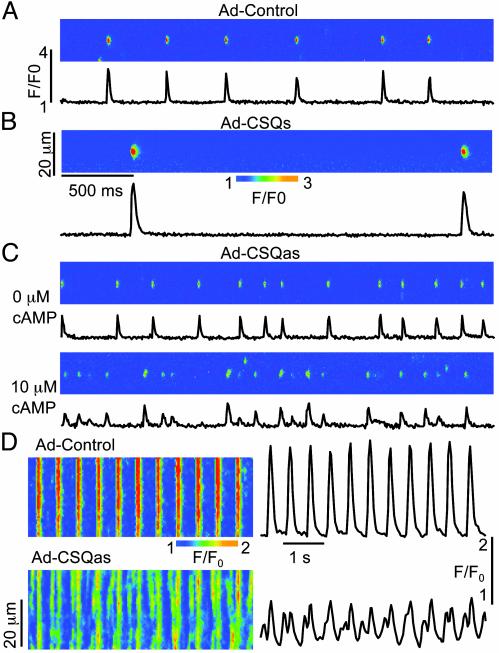
Calsequestrin is a high-capacity Ca-binding protein expressed inside the sarcoplasmic reticulum (SR), an intracellular Ca release and storage organelle in muscle. Mutations in the cardiac calsequestrin gene (CSQ2) have been linked to arrhythmias and sudden death. We have used Ca-imaging and patch-clamp methods in combination with adenoviral gene transfer strategies to explore the function of CSQ2 in adult rat heart cells. By increasing or decreasing CSQ2 levels, we showed that CSQ2 not only determines the Ca storage capacity of the SR but also positively controls the amount of Ca released from this organelle during excitation-contraction coupling. CSQ2 controls Ca release by prolonging the duration of Ca fluxes through the SR Ca-release sites. In addition, the dynamics of functional restitution of Ca-release sites after Ca discharge were prolonged when CSQ2 levels were elevated and accelerated in the presence of lowered CSQ2 protein levels. Furthermore, profound disturbances in rhythmic Ca transients in myocytes undergoing periodic electrical stimulation were observed when CSQ2 levels were reduced. We conclude that CSQ2 is a key determinant of the functional size and stability of SR Ca stores in cardiac muscle. CSQ2 appears to exert its effects by influencing the local luminal Ca concentration-dependent gating of the Ca-release channels and by acting as both a reservoir and a sink for Ca in SR. The abnormal restitution of Ca-release channels in the presence of reduced CSQ2 levels provides a plausible explanation for ventricular arrhythmia associated with mutations of CSQ2.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
