

Efficient and specific repair of sickle beta-globin RNA by trans-splicing ribozymes
- PMID: 13130139
- PMCID: PMC1370489
- DOI: 10.1261/rna.5450203
Efficient and specific repair of sickle beta-globin RNA by trans-splicing ribozymes
Abstract
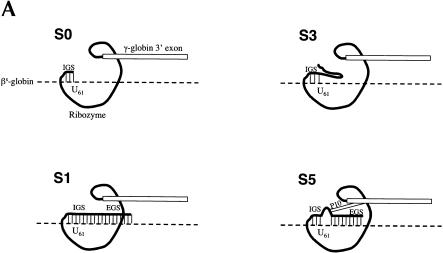
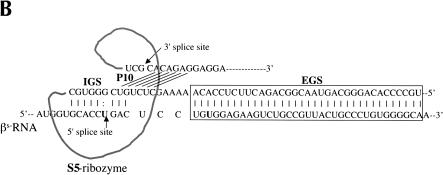
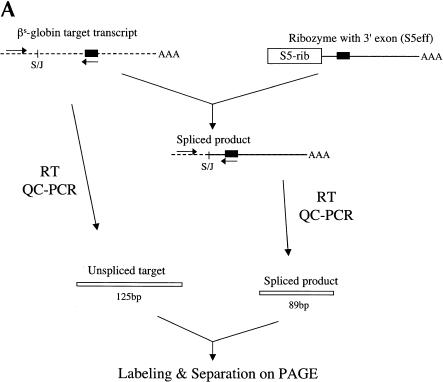
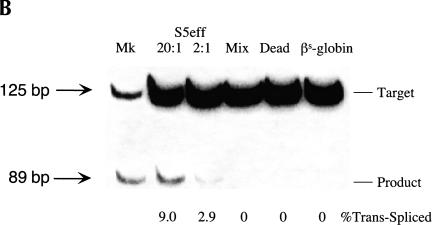
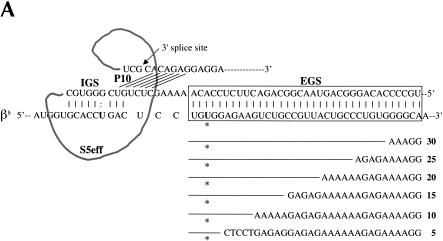
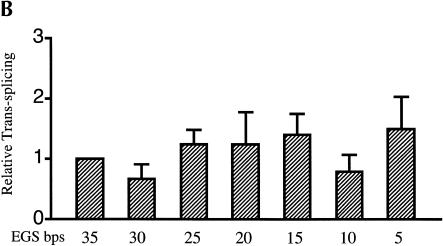
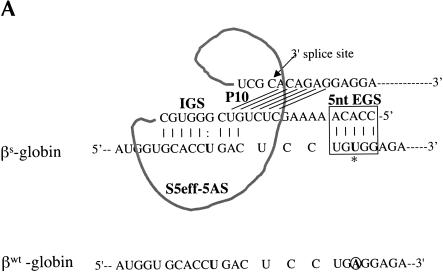
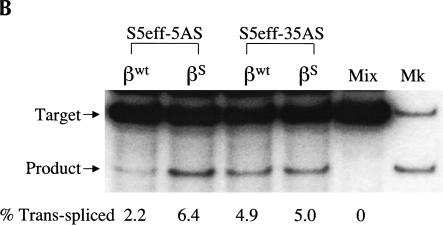
Previously we demonstrated that a group I ribozyme can perform trans-splicing to repair sickle beta-globin transcripts upon transfection of in vitro transcribed ribozyme into mammalian cells. Here, we sought to develop expression cassettes that would yield high levels of active ribozyme after gene transfer. Our initial expression constructs were designed to generate trans-slicing ribozymes identical to those used in our previous RNA transfection studies with ribozymes containing 6-nucleotide long internal guide sequences. The ribozymes expressed from these cassettes, however, were found to be unable to repair sickle beta-globin RNAs. Further experiments revealed that two additional structural elements are important for ribozyme-mediate RNA repair: the P10 interaction formed between the 5' end of the ribozyme and the beginning of the 3' exon and an additional base-pairing interaction formed between an extended guide sequence and the substrate RNA. These optimized expression cassettes yield ribozymes that are able to amend 10%-50% of the sickle beta-globin RNAs in transfected mammalian cells. Finally, a ribozyme with a 5-bp extended guide sequence preferentially reacts with sickle beta-globin RNAs over wild-type beta-globin RNAs, although the wild-type beta-globin transcript forms only a single mismatch with the ribozyme. These results demonstrate that trans-splicing ribozyme expression cassettes can be generated to yield ribozymes that can repair a clinically relevant fraction of sickle beta-globin RNAs in mammalian cells with greatly improved specificity.
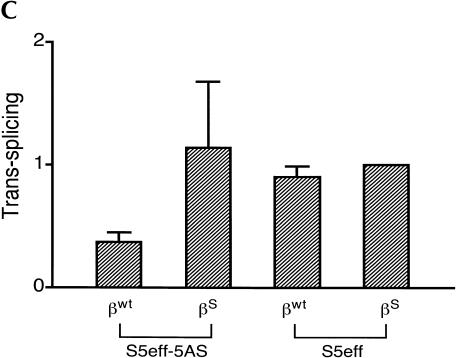
Figures

Similar articles
-
Enhancing RNA repair efficiency by combining trans-splicing ribozymes that recognize different accessible sites on a target RNA.Mol Ther. 2000 Sep;2(3):245-55. doi: 10.1006/mthe.2000.0125. Mol Ther. 2000. PMID: 10985955
-
Ribozyme-mediated repair of sickle beta-globin mRNAs in erythrocyte precursors.Science. 1998 Jun 5;280(5369):1593-6. doi: 10.1126/science.280.5369.1593. Science. 1998. PMID: 9616120
-
RNA reprogramming of alpha-mannosidase mRNA sequences in vitro by myxomycete group IC1 and IE ribozymes.FEBS J. 2006 Jun;273(12):2789-800. doi: 10.1111/j.1742-4658.2006.05295.x. FEBS J. 2006. PMID: 16817905
-
Group I Intron-Based Therapeutics Through Trans-Splicing Reaction.Prog Mol Biol Transl Sci. 2018;159:79-100. doi: 10.1016/bs.pmbts.2018.07.001. Epub 2018 Aug 9. Prog Mol Biol Transl Sci. 2018. PMID: 30340790 Review.
-
RNA reprogramming and repair based on trans-splicing group I ribozymes.N Biotechnol. 2010 Jul 31;27(3):194-203. doi: 10.1016/j.nbt.2010.02.013. Epub 2010 Feb 26. N Biotechnol. 2010. PMID: 20219714 Review.
Cited by
-
Design and Experimental Evolution of trans-Splicing Group I Intron Ribozymes.Molecules. 2017 Jan 2;22(1):75. doi: 10.3390/molecules22010075. Molecules. 2017. PMID: 28045452 Free PMC article. Review.
-
In vivo excision of a single targeted nucleotide from an mRNA by a trans excision-splicing ribozyme.RNA. 2005 Jun;11(6):897-905. doi: 10.1261/rna.2050505. Epub 2005 May 4. RNA. 2005. PMID: 15872183 Free PMC article.
-
Targeting of highly conserved Dengue virus sequences with anti-Dengue virus trans-splicing group I introns.BMC Mol Biol. 2010 Nov 15;11:84. doi: 10.1186/1471-2199-11-84. BMC Mol Biol. 2010. PMID: 21078188 Free PMC article.
-
Low selection pressure aids the evolution of cooperative ribozyme mutations in cells.J Biol Chem. 2013 Nov 15;288(46):33096-106. doi: 10.1074/jbc.M113.511469. Epub 2013 Oct 2. J Biol Chem. 2013. PMID: 24089519 Free PMC article.
-
RNA Trans-Splicing Targeting Endogenous β-Globin Pre-Messenger RNA in Human Erythroid Cells.Hum Gene Ther Methods. 2017 Apr;28(2):91-99. doi: 10.1089/hgtb.2016.077. Epub 2017 Feb 14. Hum Gene Ther Methods. 2017. PMID: 28267358 Free PMC article.
References
-
- Been, M.D. and Cech, T.R. 1986. One binding site determines sequence specificity of Tetrahymena pre-rRNA self-splicing, trans-splicing, and RNA enzyme activity. Cell 47: 207–216. - PubMed
-
- Couture, L.A. and Stinchcomb, D.T. 1996. Anti-gene therapy: The use of ribozymes to inhibit gene function. Trends Genet. 12: 510– 515. - PubMed
-
- Elbashir, S.M., Harborth, J., Lendeckel, W., Yalcin, A., Weber, K., Tuschl, T. 2001. Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature 411: 494–498. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical