

5' exon replacement and repair by spliceosome-mediated RNA trans-splicing
- PMID: 13130143
- PMCID: PMC1370493
- DOI: 10.1261/rna.5101903
5' exon replacement and repair by spliceosome-mediated RNA trans-splicing
Abstract
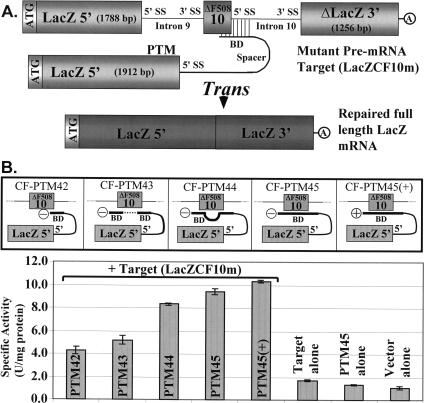
Spliceosome-mediated RNA trans-splicing (SMaRT) has been used previously to reprogram mutant endogenous CFTR and factor VIII mRNAs in human epithelial cell and tissue models and knockout mice, respectively. Those studies used 3' exon replacement (3'ER); a process in which the distal portion of RNA is reprogrammed. Here, we also show that the 5' end of mRNA can be completely rewritten by 5'ER. For proof-of-concept, and to test whether 5'ER could generate functional CFTR, we generated a mutant minigene target containing CFTR exons 10-24 (deltaF508) and a mini-intron 10, and a pretrans-splicing molecule (targeted to intron 10) containing CFTR exons 1-10 (+F508), and tested these two constructs in 293T cells for anion efflux transport. Cells cotransfected with target and PTM showed a consistent increase in anion efflux, but there was no response in control cells that received PTM or target alone. Using a LacZ reporter system to accurately quantify trans-splicing efficiency, we tested several unique PTM designs. These studies provided two important findings as follows: (1) efficient trans-splicing can be achieved by binding the PTM to different locations in the target, and (2) relatively few changes in PTM design can have a profound impact on trans-splicing activity. Tethering the PTM close to the target 3' splice site (as opposed to the donor site) and inserting an intron in the PTM coding resulted in a 65-fold enhancement of LacZ activity. These studies demonstrate that (1) SMaRT can be used to reprogram the 5' end of mRNA, and (2) efficiency can be improved substantially.
Figures




References
-
- Berget, S.M. 1995. Exon recognition in vertebrate splicing. J. Biol. Chem. 270: 2411–2414. - PubMed
-
- Chao, H., Mansfield, S.G., Bartel, R.C., Hiriyanna, S., Mitchell, L.G., Garcia-Blanco, M.A., and Walsh, C.E. 2003. Phenotype correction of hemophilia A mice by spliceosome-mediated RNA trans-splicing. Nat. Med. 9: 1015–1019. - PubMed
-
- Chowrira, B.M., Pavco, P.A., and McSwiggen, J.A. 1994. In vitro and in vivo comparison of hammerhead, hairpin, and hepatitis δ virus self-processing ribozyme cassettes. J. Biol. Chem. 269: 25856– 25864. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical