

Basilar membrane responses to two-tone and broadband stimuli
- PMID: 1354369
- PMCID: PMC3578387
- DOI: 10.1098/rstb.1992.0063
Basilar membrane responses to two-tone and broadband stimuli
Abstract
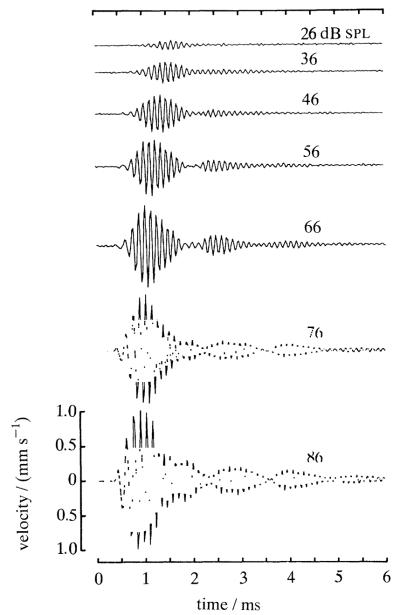
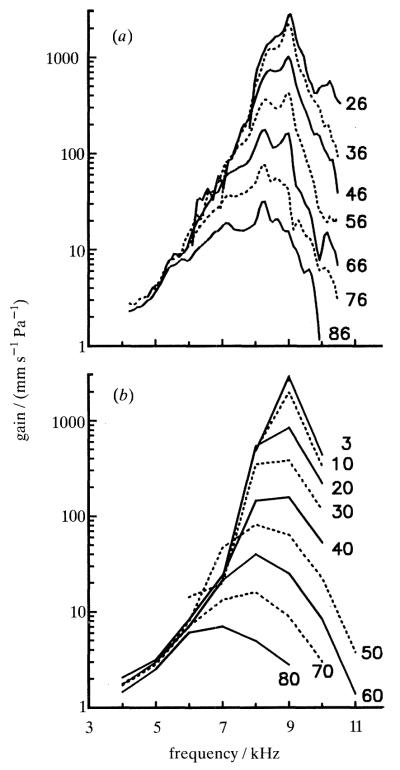
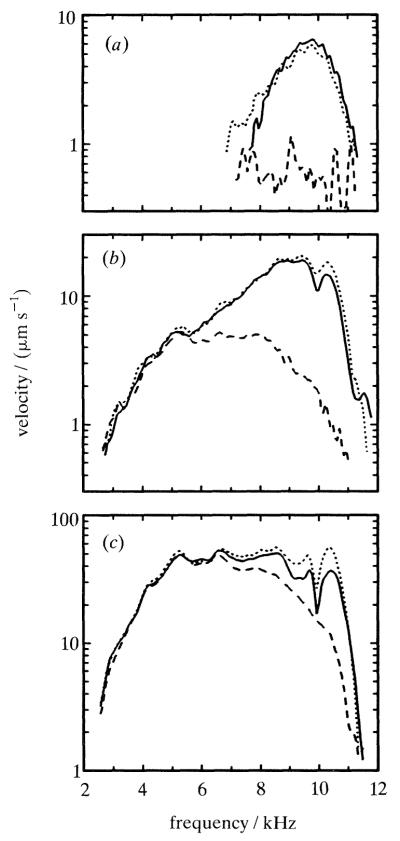
The responses to sound of mammalian cochlear neurons exhibit many nonlinearities, some of which (such as two-tone rate suppression and intermodulation distortion) are highly frequency specific, being strongly tuned to the characteristic frequency (CF) of the neuron. With the goal of establishing the cochlear origin of these auditory-nerve nonlinearities, mechanical responses to clicks and to pairs of tones were studied in relatively healthy chinchilla cochleae at a basal site of the basilar membrane with CF of 8-10 kHz. Responses were also obtained in cochleae in which hair cell receptor potentials were reduced by systemic furosemide injection. Vibrations were recorded using either the Mössbauer technique or laser Doppler-shift velocimetry. Responses to tone pairs contained intermodulation distortion products whose magnitudes as a function of stimulus frequency and intensity were comparable to those of distortion products in cochlear afferent responses. Responses to CF tones could be selectively suppressed by tones with frequency either higher or lower than CF; in most respects, mechanical two-tone suppression resembled rate suppression in cochlear afferents. Responses to clicks displayed a CF-specific compressive nonlinearity, similar to that present in responses to single tones, which could be profoundly and selectively reduced by furosemide. The present findings firmly support the hypothesis that all CF-specific nonlinearities present in the auditory nerve originate in analogous phenomena of basilar membrane vibration. However, because of their lability, it is almost certain that the mechanical nonlinearities themselves originate in outer hair cells.
Figures
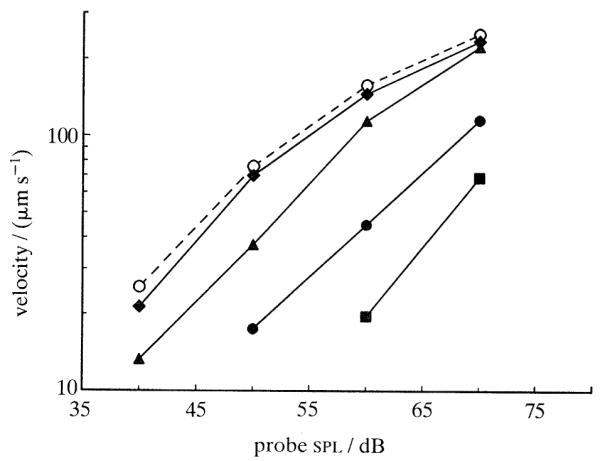
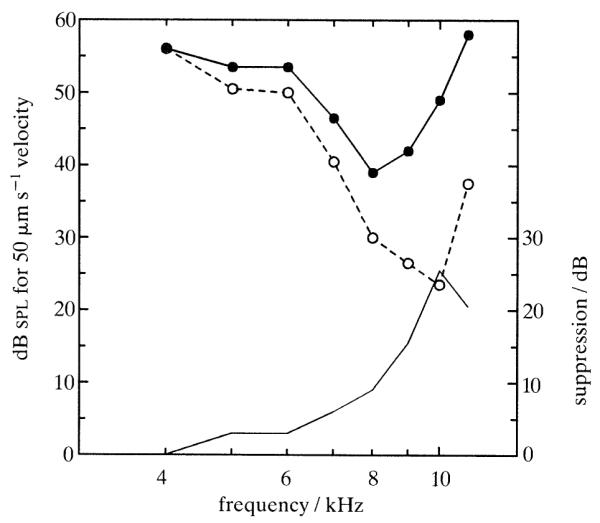
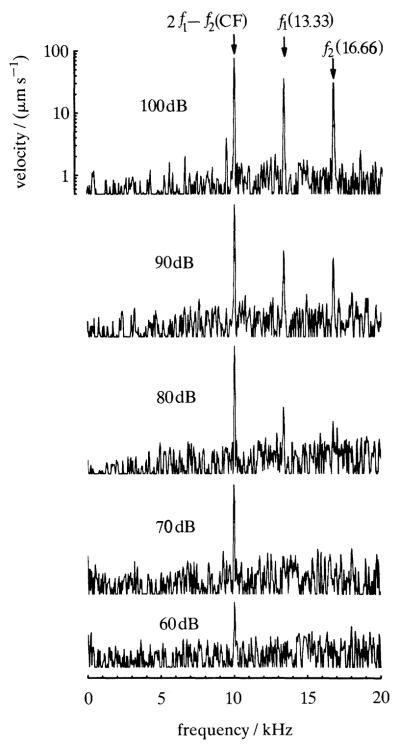
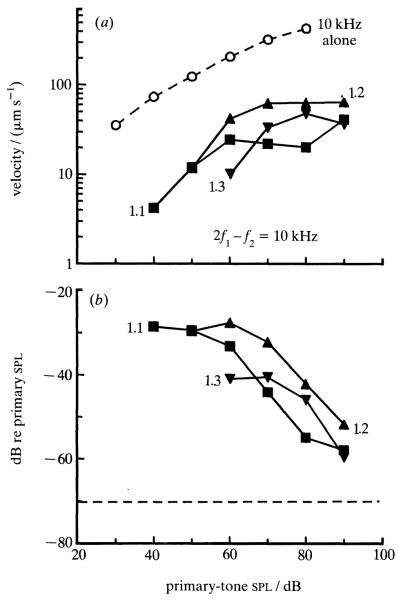
Similar articles
-
Two-tone suppression in the basilar membrane of the cochlea: mechanical basis of auditory-nerve rate suppression.J Neurophysiol. 1992 Oct;68(4):1087-99. doi: 10.1152/jn.1992.68.4.1087. J Neurophysiol. 1992. PMID: 1432070
-
Furosemide alters organ of corti mechanics: evidence for feedback of outer hair cells upon the basilar membrane.J Neurosci. 1991 Apr;11(4):1057-67. doi: 10.1523/JNEUROSCI.11-04-01057.1991. J Neurosci. 1991. PMID: 2010805 Free PMC article.
-
Wiener kernels of chinchilla auditory-nerve fibers: verification using responses to tones, clicks, and noise and comparison with basilar-membrane vibrations.J Neurophysiol. 2005 Jun;93(6):3635-48. doi: 10.1152/jn.00885.2004. Epub 2005 Jan 19. J Neurophysiol. 2005. PMID: 15659530 Free PMC article.
-
Basilar membrane motion in relation to two-tone suppression.Hear Res. 1998 Jan;115(1-2):129-42. doi: 10.1016/s0378-5955(97)00187-1. Hear Res. 1998. PMID: 9472742 Review.
-
Mechanics of the mammalian cochlea.Physiol Rev. 2001 Jul;81(3):1305-52. doi: 10.1152/physrev.2001.81.3.1305. Physiol Rev. 2001. PMID: 11427697 Free PMC article. Review.
Cited by
-
Multiple sounds degrade the frequency representation in monkey inferior colliculus.Eur J Neurosci. 2022 Jan;55(2):528-548. doi: 10.1111/ejn.15545. Epub 2021 Dec 30. Eur J Neurosci. 2022. PMID: 34844286 Free PMC article.
-
Medial olivocochlear efferent inhibition of basilar-membrane responses to clicks: evidence for two modes of cochlear mechanical excitation.J Acoust Soc Am. 2008 Aug;124(2):1080-92. doi: 10.1121/1.2949435. J Acoust Soc Am. 2008. PMID: 18681598 Free PMC article.
-
A Level-Adjusted Cochlear Frequency-to-Place Map for Estimating Tonotopic Frequency Mismatch With a Cochlear Implant.Ear Hear. 2025 Jul-Aug 01;46(4):963-975. doi: 10.1097/AUD.0000000000001641. Epub 2025 Feb 11. Ear Hear. 2025. PMID: 39930567
-
Basilar-membrane responses to clicks at the base of the chinchilla cochlea.J Acoust Soc Am. 1998 Apr;103(4):1972-89. doi: 10.1121/1.421377. J Acoust Soc Am. 1998. PMID: 9566320 Free PMC article.
-
Responses to sound of the basilar membrane of the mammalian cochlea.Curr Opin Neurobiol. 1992 Aug;2(4):449-56. doi: 10.1016/0959-4388(92)90179-o. Curr Opin Neurobiol. 1992. PMID: 1525542 Free PMC article. Review.
References
-
- Brownell WE, Bader CR, Bertrand D, de Ribaupierre Y. Evoked mechanical responses of isolated outer hair cells. Science, Wash. 1985;227:194–196. - PubMed
-
- Brundin L, Flock A, Canlon B. Sound-induced motility of isolated cochlear outer hair cells is frequency-specific. Nature, Lond. 1989;342:814–816. - PubMed
-
- Buunen TJF, Rhode WS. Responses of fibers in the cat’s auditory nerve to the cubic difference tone. J. acoust. Soc. Am. 1978;64:772–781. - PubMed
-
- Cody AR, Mountain DC. Low-frequency responses of inner hair cells: evidence for a mechanical origin of peak splitting. Hear. Res. 1989;41:89–100. - PubMed
-
- Costalupes JA, Rich NC, Ruggero MA. Effects of excitatory and non-excitatory suppressor tones on two-tone rate suppression in auditory nerve fibers. Hear. Res. 1987;26:155–164. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
