

Identification of novel recognition motifs and regulatory targets for the yeast actin-regulating kinase Prk1p
- PMID: 13679512
- PMCID: PMC284791
- DOI: 10.1091/mbc.e03-06-0362
Identification of novel recognition motifs and regulatory targets for the yeast actin-regulating kinase Prk1p
Abstract
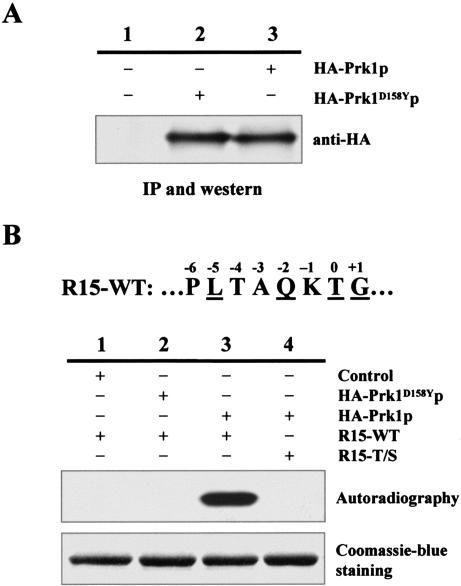
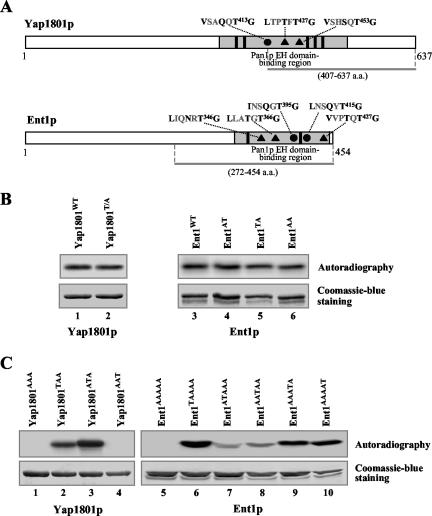
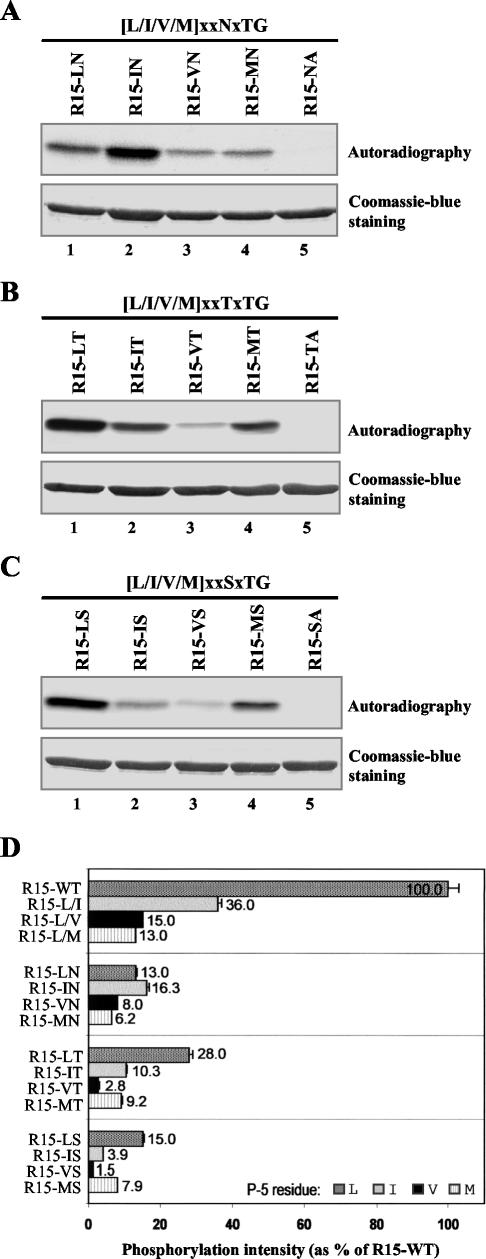
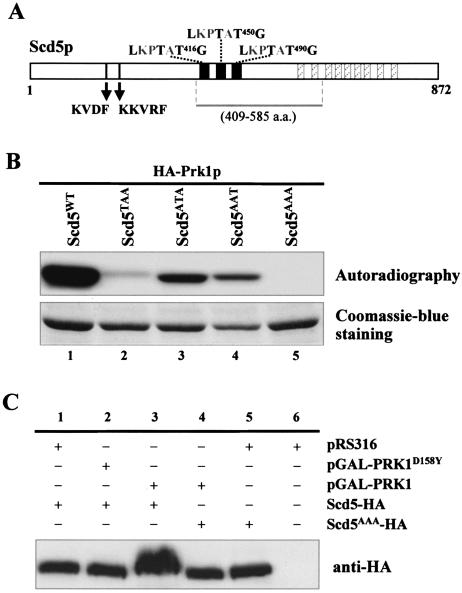
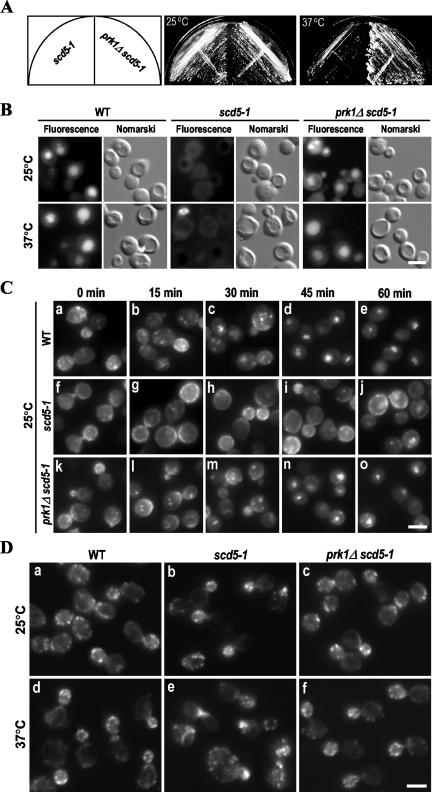
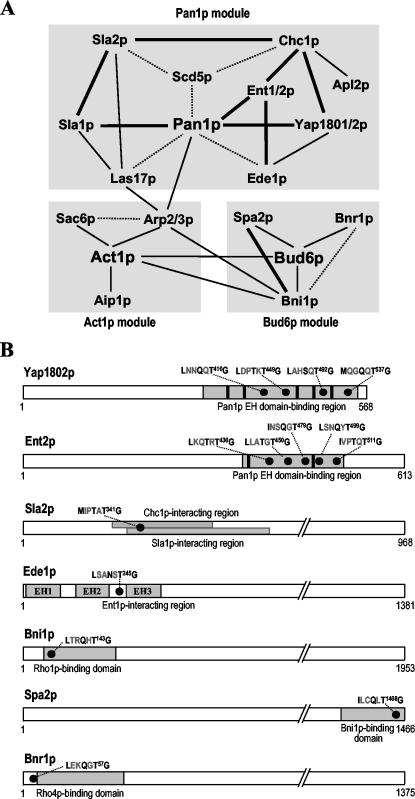
Prk1p is a serine/threonine kinase involved in the regulation of the actin cytoskeleton organization in the yeast Saccharomyces cerevisiae. Previously, we have identified LxxQxTG as the phosphorylation site of Prk1p. In this report, the recognition sequence for Prk1p is investigated more thoroughly. It is found that the presence of a hydrophobic residue at the position of P-5 is necessary for Prk1p phosphorylation and L, I, V, and M are all able to confer the phosphorylation at various efficiencies. The residue flexibility at P-2 has also been identified to include Q, N, T, and S. A homology-based three-dimensional model of the kinase domain of Prk1p provided some structural interpretations for these substrate specificities. The characterization of the [L/I/V/M]xx[Q/N/T/S]xTG motif led to the identification of a spectrum of potential targets for Prk1p from yeast genome. One of them, Scd5p, which contains three LxxTxTG motifs and is previously known to be important for endocytosis and actin organization, has been chosen to demonstrate its relationship with Prk1p. Phosphorylation of Scd5p by Prk1p at the three LxxTxTG motifs could be detected in vitro and in vivo, and deletion of PRK1 suppressed the defects in actin cytoskeleton and endocytosis in one of the scd5 mutants. These results allowed us to conclude that Scd5p is likely another regulatory target of Prk1p.
Figures


Similar articles
-
Prk1p.Int J Biochem Cell Biol. 2005 Jan;37(1):48-53. doi: 10.1016/j.biocel.2004.03.010. Int J Biochem Cell Biol. 2005. PMID: 15381149 Review.
-
Regulation of yeast actin cytoskeleton-regulatory complex Pan1p/Sla1p/End3p by serine/threonine kinase Prk1p.Mol Biol Cell. 2001 Dec;12(12):3759-72. doi: 10.1091/mbc.12.12.3759. Mol Biol Cell. 2001. PMID: 11739778 Free PMC article.
-
Regulation of the actin cytoskeleton organization in yeast by a novel serine/threonine kinase Prk1p.J Cell Biol. 1999 Jan 11;144(1):71-82. doi: 10.1083/jcb.144.1.71. J Cell Biol. 1999. PMID: 9885245 Free PMC article.
-
Novel protein kinases Ark1p and Prk1p associate with and regulate the cortical actin cytoskeleton in budding yeast.J Cell Biol. 1999 Mar 22;144(6):1203-18. doi: 10.1083/jcb.144.6.1203. J Cell Biol. 1999. PMID: 10087264 Free PMC article.
-
Pan1p: an actin director of endocytosis in yeast.Int J Biochem Cell Biol. 2007;39(10):1760-4. doi: 10.1016/j.biocel.2006.12.001. Epub 2006 Dec 21. Int J Biochem Cell Biol. 2007. PMID: 17303466 Review.
Cited by
-
Deciphering protein kinase specificity through large-scale analysis of yeast phosphorylation site motifs.Sci Signal. 2010 Feb 16;3(109):ra12. doi: 10.1126/scisignal.2000482. Sci Signal. 2010. PMID: 20159853 Free PMC article.
-
Yeast Eps15-like endocytic protein Pan1p regulates the interaction between endocytic vesicles, endosomes and the actin cytoskeleton.Elife. 2016 Feb 25;5:e10276. doi: 10.7554/eLife.10276. Elife. 2016. PMID: 26914139 Free PMC article.
-
Actin and endocytosis in budding yeast.Genetics. 2015 Feb;199(2):315-58. doi: 10.1534/genetics.112.145540. Genetics. 2015. PMID: 25657349 Free PMC article. Review.
-
Cortical recruitment and nuclear-cytoplasmic shuttling of Scd5p, a protein phosphatase-1-targeting protein involved in actin organization and endocytosis.Mol Biol Cell. 2006 Jan;17(1):251-62. doi: 10.1091/mbc.e05-10-0936. Epub 2005 Oct 26. Mol Biol Cell. 2006. PMID: 16251346 Free PMC article.
-
Cdc28-Cln3 phosphorylation of Sla1 regulates actin patch dynamics in different modes of fungal growth.Mol Biol Cell. 2012 Sep;23(17):3485-97. doi: 10.1091/mbc.E12-03-0231. Epub 2012 Jul 11. Mol Biol Cell. 2012. PMID: 22787279 Free PMC article.
References
-
- Adams, A.E., and Pringle, J.R. (1991). Staining of actin with fluorochrome-conjugated phalloidin. Methods Enzymol. 194, 729–731. - PubMed
-
- Aguilar, R.C., Watson, H.A., and Wendland, B. (2003). The yeast Epsin Ent1 is recruited to membranes through multiple independent interactions. J. Biol. Chem. 278, 10737–10743. - PubMed
-
- Chang, F., and Peter, M. (2003). Yeasts make their mark. Nat. Cell Biol. 5, 294–299. - PubMed
-
- Chang, J.S., Henry, K., Wolf, B.L., Geli, M., and Lemmon, S.K. (2002). Protein phosphatase-1 binding to scd5p is important for regulation of actin organization and endocytosis in yeast. J. Biol. Chem. 277, 48002–48008. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
