

The Drosophila kinesin-like protein KLP67A is essential for mitotic and male meiotic spindle assembly
- PMID: 13679514
- PMCID: PMC307533
- DOI: 10.1091/mbc.e03-05-0342
The Drosophila kinesin-like protein KLP67A is essential for mitotic and male meiotic spindle assembly
Abstract
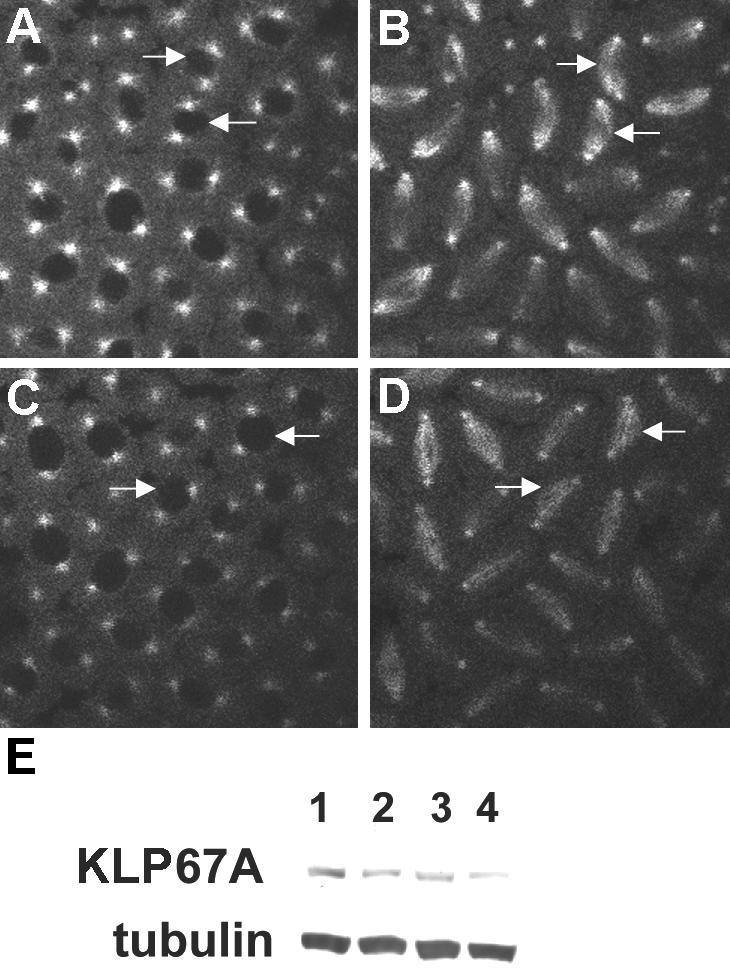
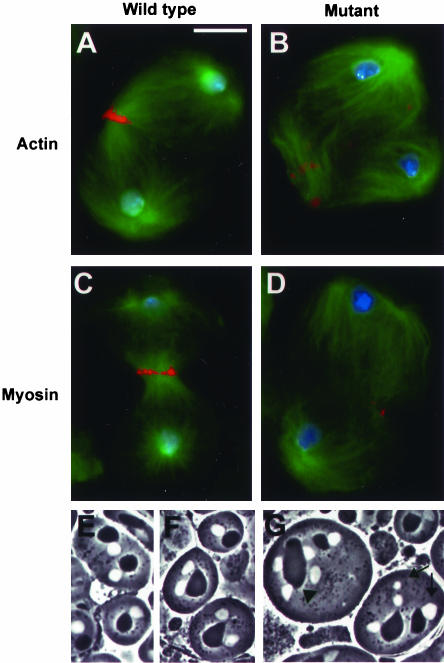
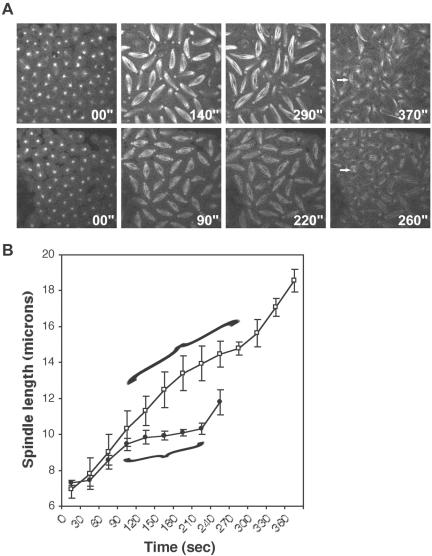
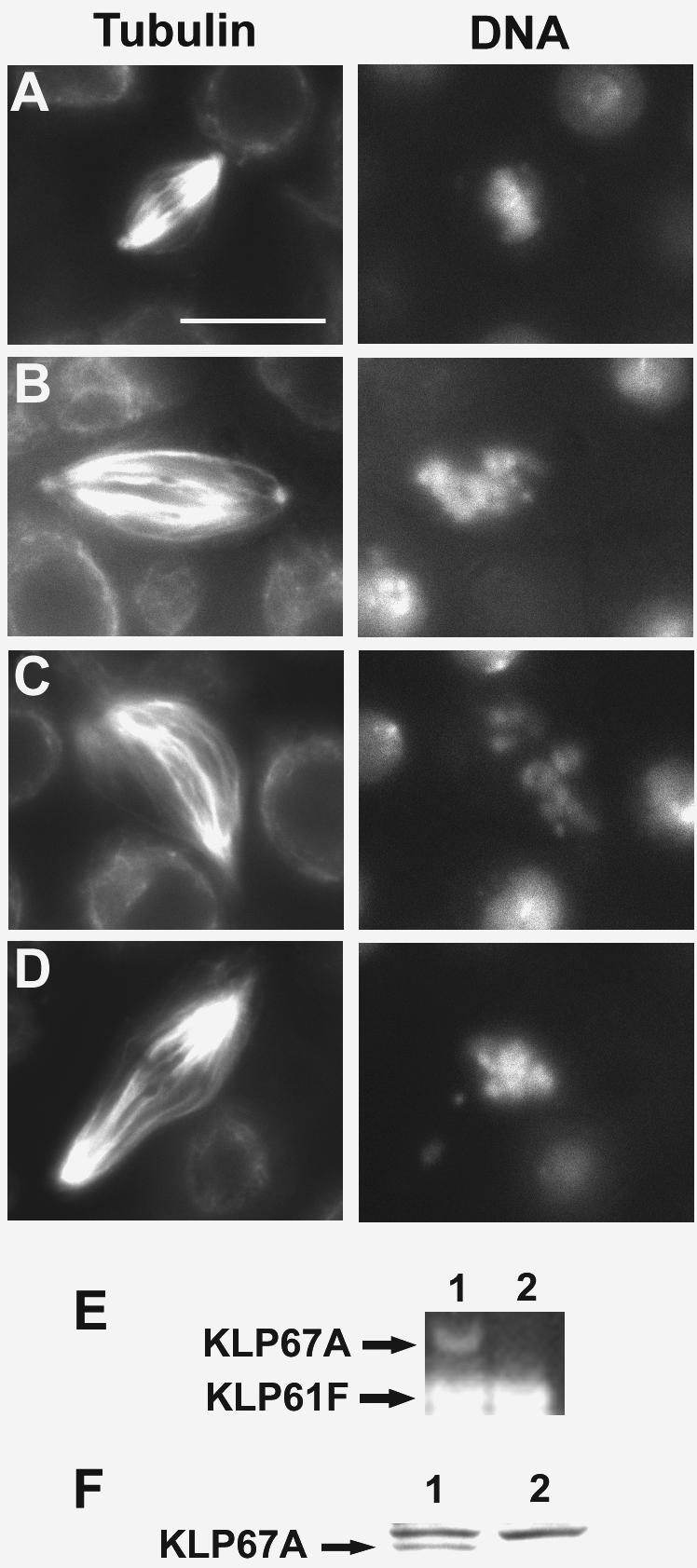
We have performed a mutational analysis together with RNA interference to determine the role of the kinesin-like protein KLP67A in Drosophila cell division. During both mitosis and male meiosis, Klp67A mutations cause an increase in MT length and disrupt discrete aspects of spindle assembly, as well as cytokinesis. Mutant cells exhibit greatly enlarged metaphase spindle as a result of excessive MT polymerization. The analysis of both living and fixed cells also shows perturbations in centrosome separation, chromosome segregation, and central spindle assembly. These data demonstrate that the MT plus end-directed motor KLP67A is essential for spindle assembly during mitosis and male meiosis and suggest that the regulation of MT plus-end polymerization is a key determinant of spindle architecture throughout cell division.
Figures



References
-
- Basto, R., Gomes, R., and Karess, R.E. (2000). Rough Deal and Zw10 are required for the metaphase checkpoint in Drosophila. Nat. Cell Biol. 2, 939–943. - PubMed
-
- Cassimeris, L. (1999). Accessory protein regulation of microtubule dynamics throughout the cell cycle. Curr. Opin. Cell Biol. 11, 134–141. - PubMed
-
- Cenci, G., Bonaccorsi, S., Pisano, C., Verni, F., and Gatti, M. (1994). Chromatin and microtubule organization during premeiotic meiotic, and early postmeiotic stages of Drosophila melanogaster spermatogenesis. J. Cell Sci. 107, 3521–3534. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
