

Insulin-induced phosphorylation of FKHR (Foxo1) targets to proteasomal degradation
- PMID: 13679577
- PMCID: PMC208749
- DOI: 10.1073/pnas.1934283100
Insulin-induced phosphorylation of FKHR (Foxo1) targets to proteasomal degradation
Abstract
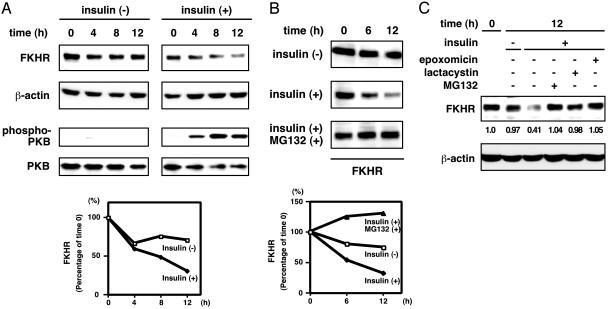
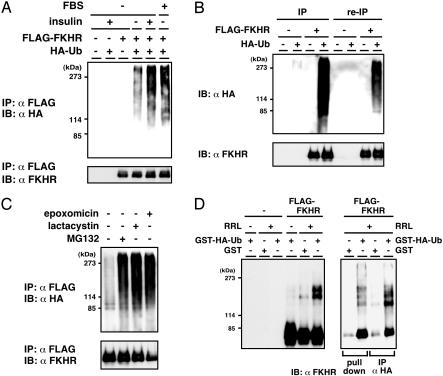
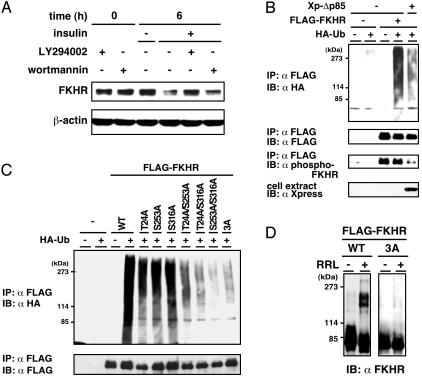
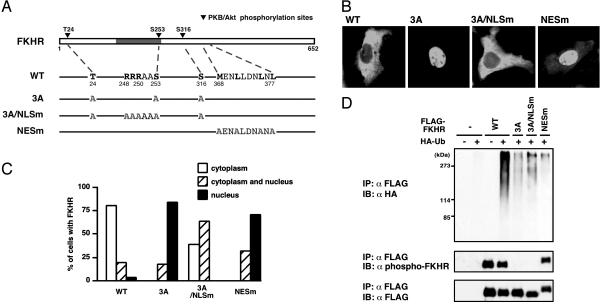
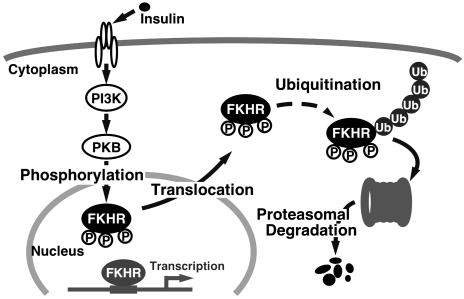
Forkhead transcription factor FKHR (Foxo1) is a key regulator of glucose homeostasis, cell-cycle progression, and apoptosis. It has been shown that FKHR is phosphorylated via insulin or growth factor signaling cascades, resulting in its cytoplasmic retention and the repression of target gene expression. Here, we investigate the fate of FKHR after cells are stimulated by insulin. We show that insulin treatment decreases endogenous FKHR proteins in HepG2 cells, which is inhibited by proteasome inhibitors. FKHR is ubiquitinated in vivo and in vitro, and insulin enhances the ubiquitination in the cells. In addition, the signal to FKHR degradation from insulin is mediated by the phosphatidylinositol 3-kinase pathway, and the mutation of FKHR at the serine or threonine residues phosphorylated by protein kinase B, a downstream target of phosphatidylinositol 3-kinase, inhibits the ubiquitination in vivo and in vitro. Finally, efficient ubiquitination of FKHR requires both phosphorylation and cytoplasmic retention in the cells. These results demonstrate that the insulin-induced phosphorylation of FKHR leads to the multistep negative regulation, not only by the nuclear exclusion but also the ubiquitination-mediated degradation.
Figures
References
-
- Kandel, E. S. & Hay, N. (1999) Exp. Cell Res. 253, 210–229. - PubMed
-
- Kido, Y., Nakae, J. & Accili, D. (2001) J. Clin. Endocrinol. Metab. 86, 972–979. - PubMed
-
- Burgering, B. M. & Kops, G. J. (2002) Trends Biochem. Sci. 27, 352–360. - PubMed
-
- Birkenkamp, K. U. & Coffer, P. J. (2003) Biochem. Soc. Trans. 31, 292–297. - PubMed
-
- Brunet, A., Bonni, A., Zigmond, M. J., Lin, M. Z., Juo, P., Hu, L. S., Anderson, M. J., Arden, K. C., Blenis, J. & Greenberg, M. E. (1999) Cell 96, 857–868. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
