

Electron microscopic evidence for nucleation and growth of 3D acetylcholine receptor microcrystals in structured lipid-detergent matrices
- PMID: 13679581
- PMCID: PMC208753
- DOI: 10.1073/pnas.1834451100
Electron microscopic evidence for nucleation and growth of 3D acetylcholine receptor microcrystals in structured lipid-detergent matrices
Abstract
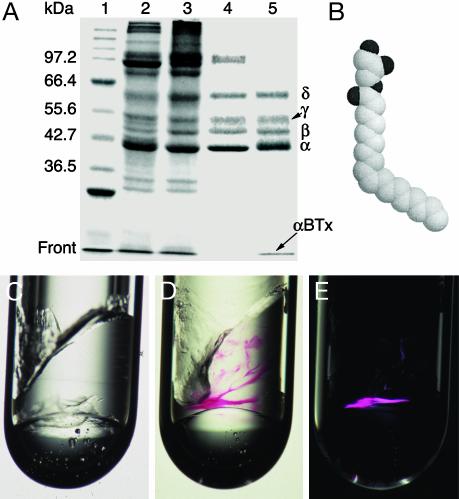
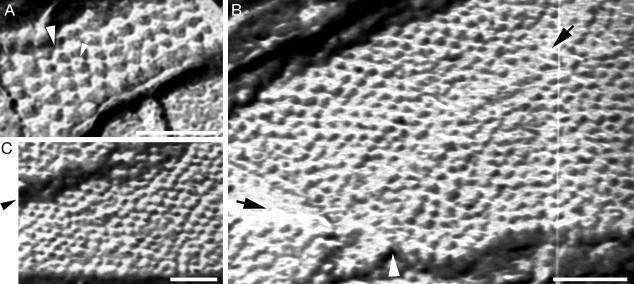
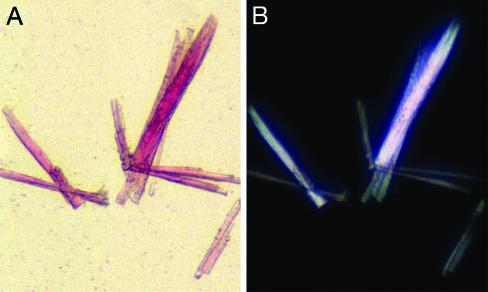
Nicotinic acetylcholine receptors (AChRs) belong to a superfamily of oligomeric proteins that transduce electric signals across the cell membrane on binding of neurotransmitters. These receptors harbor a large extracellular ligand-binding domain directly linked to an ion-conducting channel-forming domain that spans the cell membrane 20 times and considerably extends into the cytoplasm. Thus far, none of these receptor channels has been crystallized in three dimensions. The crystallization of the AChR from Torpedo marmorata electric organs is challenged here in lipidic-detergent matrices. Detergent-soluble AChR complexed with alpha-bungarotoxin (alphaBTx), a polypeptidic competitive antagonist, was purified. The AChR-alphaBTx complex was reconstituted in a lipidic matrix composed of monoolein bilayers that are structured in three dimensions. The alphaBTx was conjugated to a photo-stable fluorophore, enabling us to monitor the physical behavior of the receptor-toxin complex in the lipidic matrix under light stereomicroscope, and to freeze fracture regions containing the receptor-toxin complex for visualization under a transmission electron microscope. Conditions were established for forming 2D receptor-toxin lattices that are stacked in the third dimension. 3D AChR nanocrystals were thereby grown inside the highly viscous lipidic 3D matrix. Slow emulsification of the lipidic matrix converted these nanocrystals into 3D elongated thin crystal plates of micrometer size. The latter are stable in detergent-containing aqueous solutions and can currently be used for seeding and epitaxial growth, en route to crystals of appropriate dimensions for x-ray diffraction studies.
Figures


References
-
- Rosenbusch, J. P. (2001) J. Struct. Biol. 136, 144–157. - PubMed
-
- Corringer, P. J., Le Novere, N. & Changeux, J. P. (2000) Annu. Rev. Pharmacol. Toxicol. 40, 431–458. - PubMed
-
- Karlin, A. (2002) Nat. Rev. Neurosci. 3, 102–114. - PubMed
-
- Engel, A. G., Ohno, K. & Sine, S. M. (2003) Nat. Rev. Neurosci. 4, 339–352. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
