

Supercoiling and denaturation in Gal repressor/heat unstable nucleoid protein (HU)-mediated DNA looping
- PMID: 14500788
- PMCID: PMC208764
- DOI: 10.1073/pnas.2034851100
Supercoiling and denaturation in Gal repressor/heat unstable nucleoid protein (HU)-mediated DNA looping
Abstract
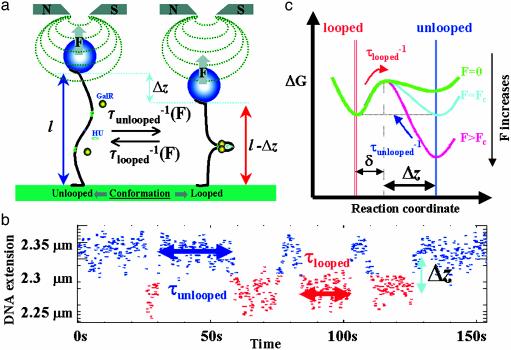
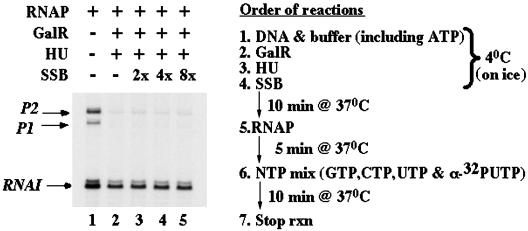
The overall topology of DNA profoundly influences the regulation of transcription and is determined by DNA flexibility as well as the binding of proteins that induce DNA torsion, distortion, and/or looping. Gal repressor (GalR) is thought to repress transcription from the two promoters of the gal operon of Escherichia coli by forming a DNA loop of approximately 40 nm of DNA that encompasses the promoters. Associated evidence of a topological regulatory mechanism of the transcription repression is the requirement for a supercoiled DNA template and the histone-like heat unstable nucleoid protein (HU). By using single-molecule manipulations to generate and finely tune tension in DNA molecules, we directly detected GalR/HU-mediated DNA looping and characterized its kinetics, thermodynamics, and supercoiling dependence. The factors required for gal DNA looping in single-molecule experiments (HU, GalR and DNA supercoiling) correspond exactly to those necessary for gal repression observed both in vitro and in vivo. Our single-molecule experiments revealed that negatively supercoiled DNA, under slight tension, denatured to facilitate GalR/HU-mediated DNA loop formation. Such topological intermediates may operate similarly in other multiprotein complexes of transcription, replication, and recombination.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
