

Stabilization of a metastable state of Torpedo californica acetylcholinesterase by chemical chaperones
- PMID: 14500892
- PMCID: PMC2366936
- DOI: 10.1110/ps.03110703
Stabilization of a metastable state of Torpedo californica acetylcholinesterase by chemical chaperones
Abstract
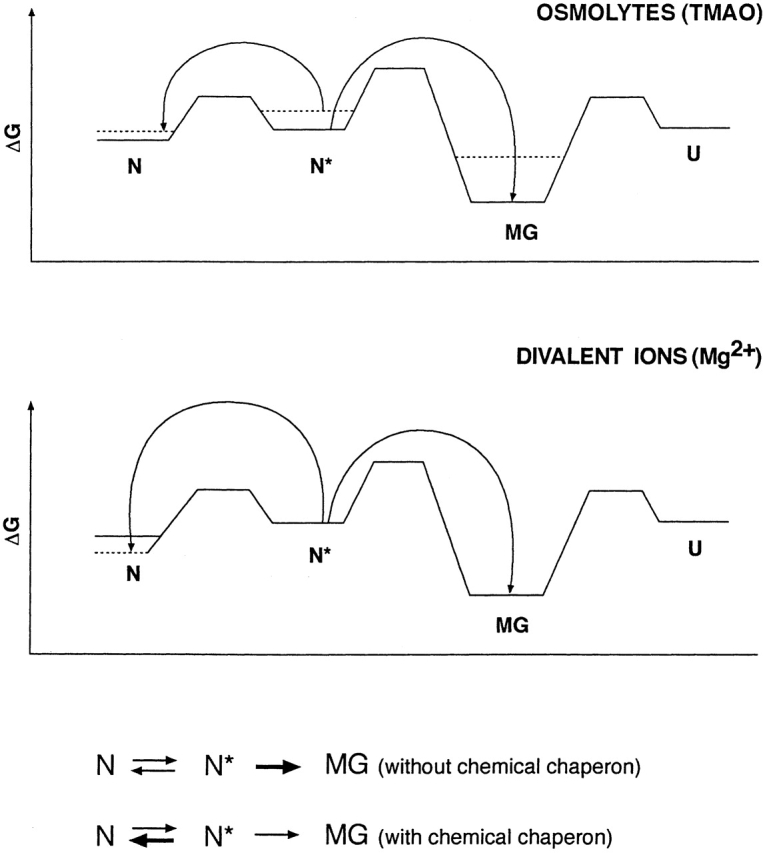
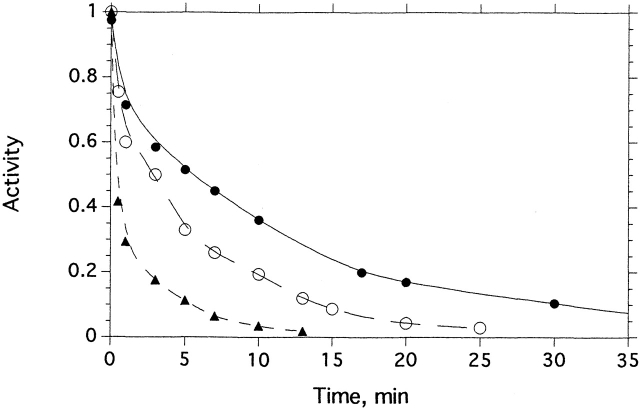
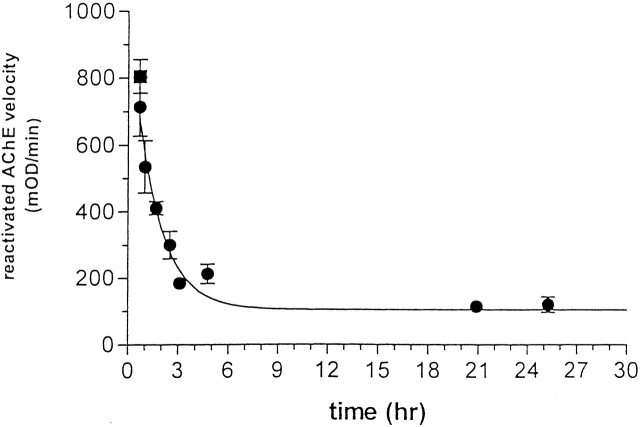
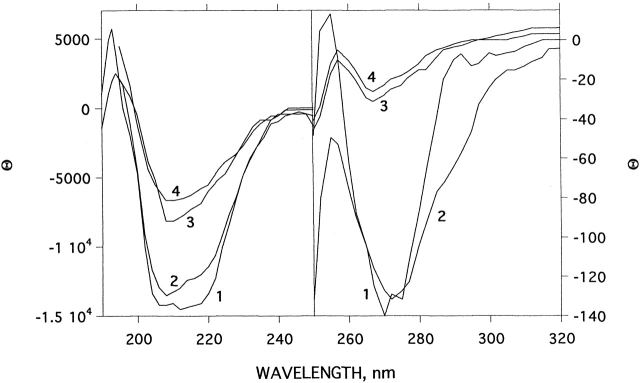
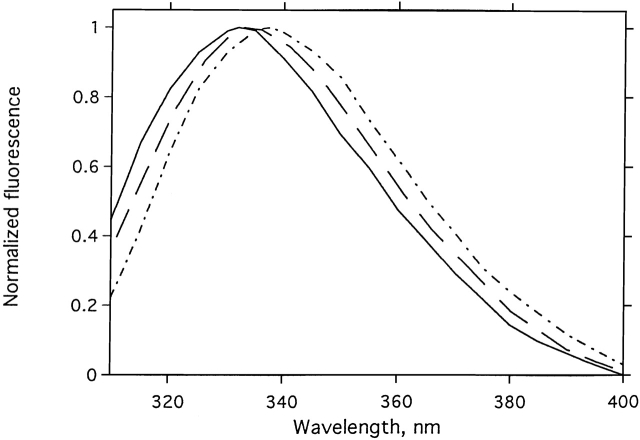
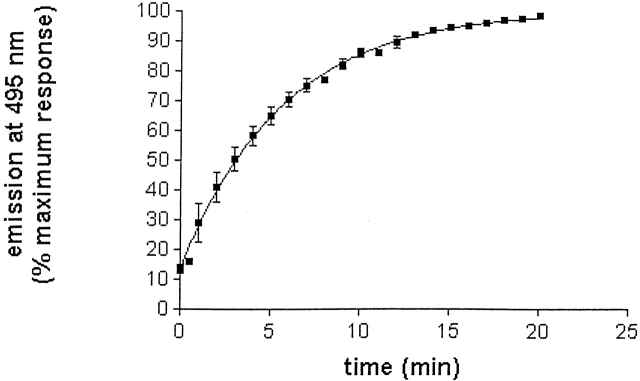
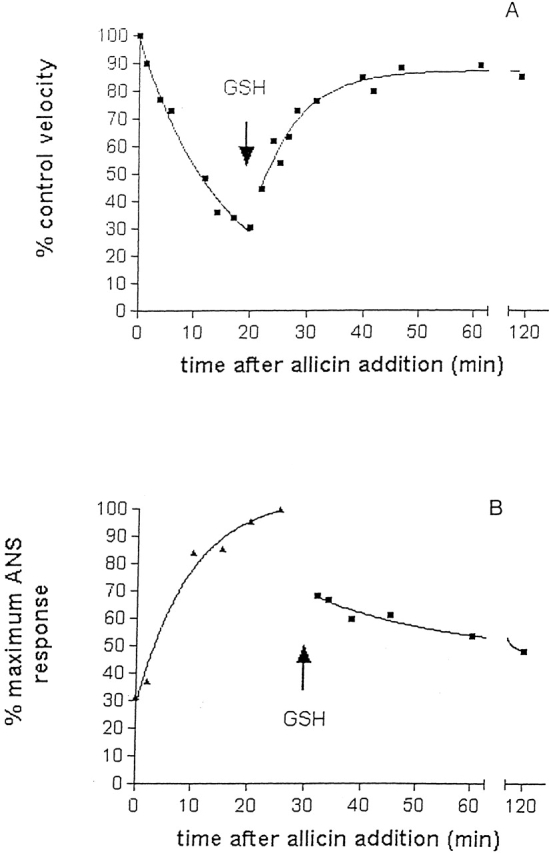
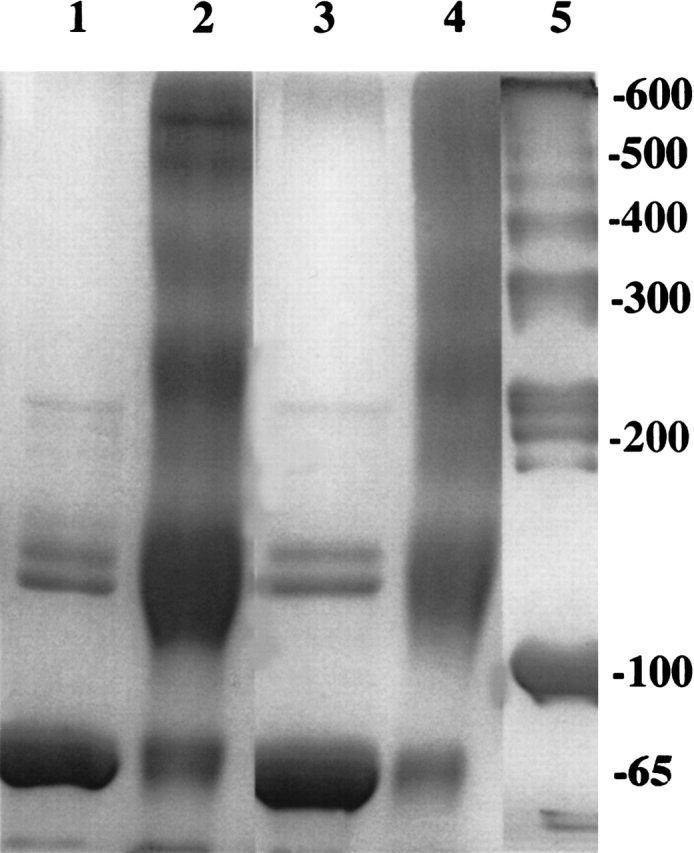
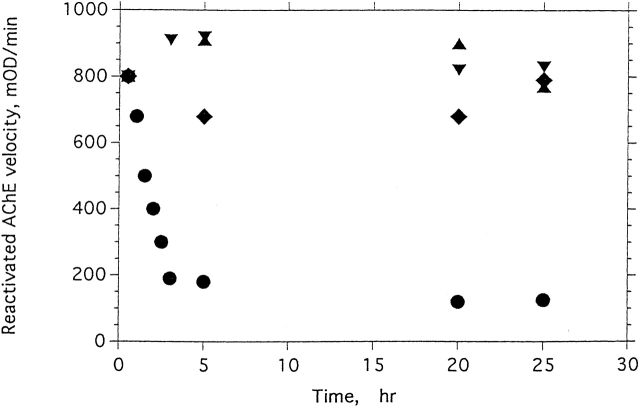
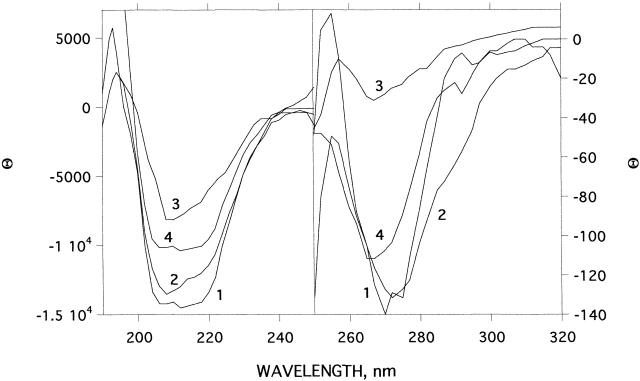
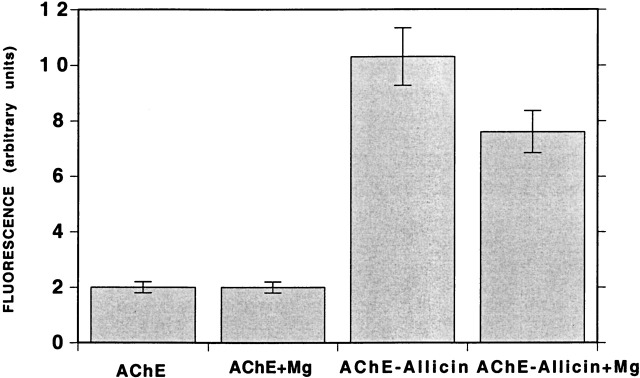
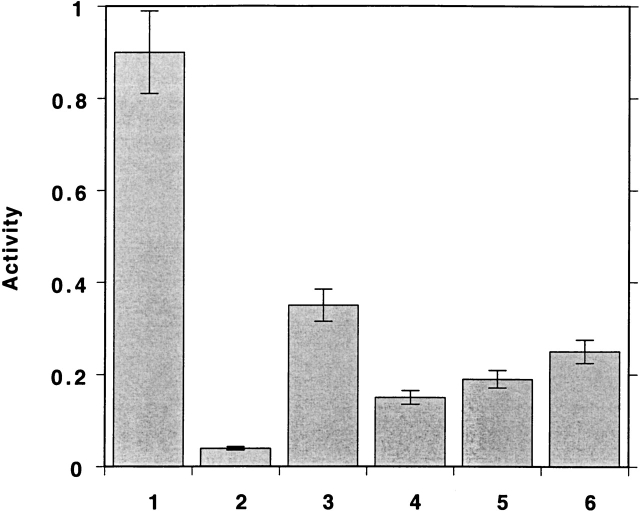
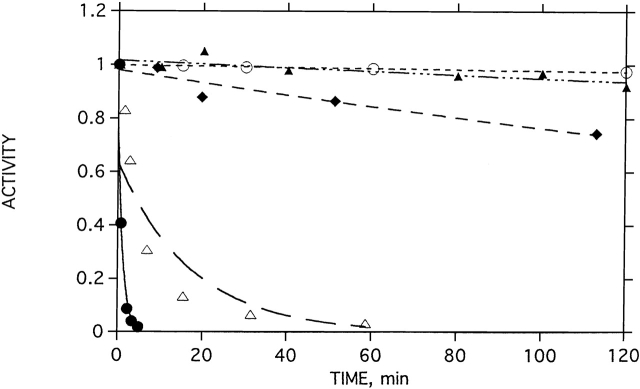
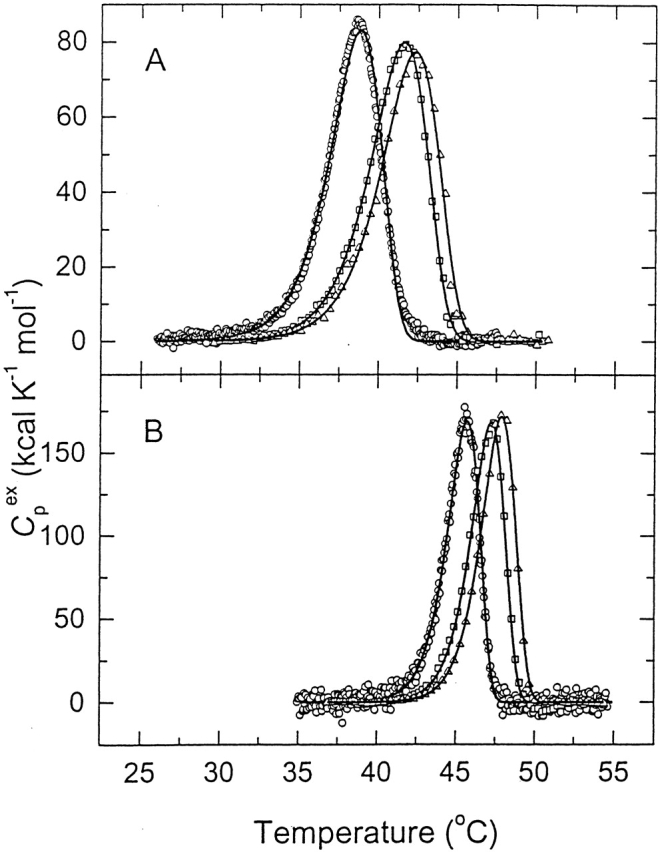
Chemical modification of Torpedo californica acetylcholinesterase by the natural thiosulfinate allicin produces an inactive enzyme through reaction with the buried cysteine Cys 231. Optical spectroscopy shows that the modified enzyme is "native-like," and inactivation can be reversed by exposure to reduced glutathione. The allicin-modified enzyme is, however, metastable, and is converted spontaneously and irreversibly, at room temperature, with t(1/2) approximately 100 min, to a stable, partially unfolded state with the physicochemical characteristics of a molten globule. Osmolytes, including trimethylamine-N-oxide, glycerol, and sucrose, and the divalent cations, Ca(2+), Mg(2+), and Mn(2+) can prevent this transition of the native-like state for >24 h at room temperature. Trimethylamine-N-oxide and Mg(2+) can also stabilize the native enzyme, with only slight inactivation being observed over several hours at 39 degrees C, whereas in their absence it is totally inactivated within 5 min. The stabilizing effects of the osmolytes can be explained by their differential interaction with the native and native-like states, resulting in a shift of equilibrium toward the native state. The stabilizing effects of the divalent cations can be ascribed to direct stabilization of the native state, as supported by differential scanning calorimetry.
Figures
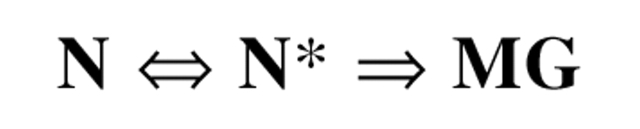
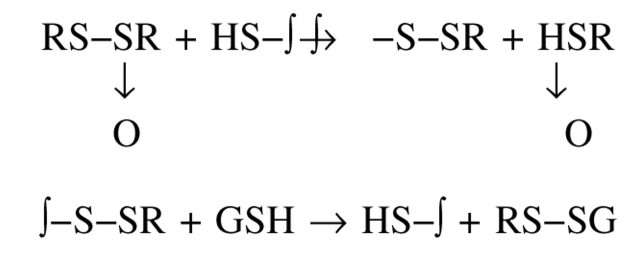
References
-
- Ananthan, J., Goldberg, A.L., and Voellmy, R. 1986. Abnormal proteins serve as eukaryotic stress signals and trigger the activation of heat shock genes. Science 232 522–524. - PubMed
-
- Arai, M. and Kuwajima, K. 2000. Role of the molten globule state in protein folding. Adv. Protein Chem. 53 209–282. - PubMed
-
- Baskakov, I.V., Legname, G., Prusiner, S.B., and Cohen, F.E. 2001. Folding of prion protein to its native α-helical conformation is under kinetic control. J. Biol. Chem. 276 19687–19690. - PubMed
-
- Bolen, D.W. and Baskakov, I.V. 2001. The osmophobic effect: Natural selection of a thermodynamic force in protein folding. J. Mol. Biol. 310 955–963. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
