

doi: 10.1073/pnas.2033520100.
Epub 2003 Sep 19.
Redefining cholesterol's role in the mechanism of the cholesterol-dependent cytolysins
Affiliations
- PMID: 14500900
- PMCID: PMC208754
- DOI: 10.1073/pnas.2033520100
Item in Clipboard
Redefining cholesterol's role in the mechanism of the cholesterol-dependent cytolysins
Proc Natl Acad Sci U S A.
.
Abstract
The cholesterol-dependent cytolysins (CDCs) constitute a large family of pore-forming toxins that function exclusively on cholesterol-containing membranes. A detailed analysis of the various stages in the cytolytic mechanism of three members of the CDC family revealed that significant depletion of cholesterol from the erythrocyte membrane stalls these toxins in the prepore complex. Therefore, the depletion of membrane cholesterol prevents the insertion of the transmembrane beta-barrel and pore formation. These unprecedented findings provide a paradigm for the involvement of cholesterol in the CDC cytolytic mechanism and that of other pore-forming toxins whose activity is enhanced by the presence of membrane cholesterol.
Figures
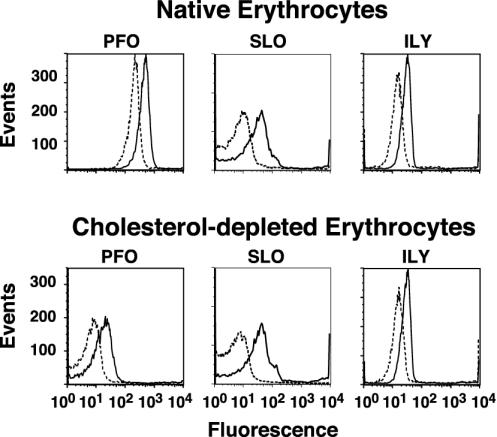
Binding to native hRBCs and cholesterol-depleted hRBCs. Binding of fluorescently labeled derivatives of PFOA215C, SLOG128C, and ILYS57C to hRBCs was determined by flow cytometry. The fluorescent-labeled toxin (4.5 pmol) was mixed with an equimolar amount of unmodified toxin, incubated with 1.5 × 106 hRBCs at 4°C for 30 min, and then analyzed by flow cytometry. We counted 104 cells per sample. Toxin binding to native (outlined peak) and cholesterol-depleted (shaded peak) hRBCs is shown for all three toxins.
Oligomerization measured by FRET and flow cytometry. Equimolar amounts (4.5 pmol each) of donor-labeled toxin (Alexa Fluor-labeled toxin) and either unlabeled (D + U, solid line) or acceptor-labeled (D + A, dashed line) toxin (tetramethylrhodamine-labeled toxin) were incubated with 1.5 × 106 hRBCs at 4°C for 30 min and then analyzed by flow cytometry. 104 cells per sample were counted. The fluorescence intensity of the donor fluorophore is shown for all three toxins on native and cholesterol-depleted hRBCs in the presence of unlabeled (D + U) or acceptor-labeled toxin (D + A). FRET between donor- and acceptor-labeled toxin is observed as a decrease in the fluorescence of the donor when the unlabeled toxin is replaced with acceptor-labeled toxin (compare D + U and D + A).
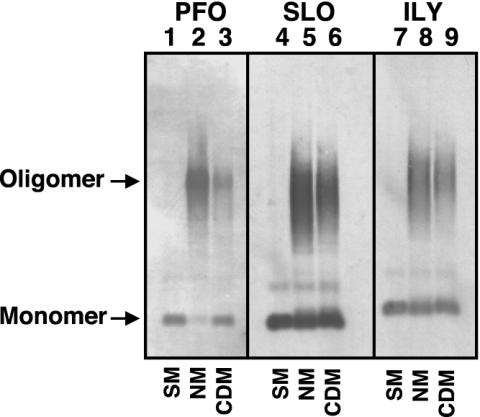
Oligomer formation on native and cholesterol-depleted hRBCs determined by SDS/AGE. The formation of oligomers of PFO, SLO, and ILY was determined by SDS-agarose electrophoresis. Each toxin (9 pmol) was incubated alone (SM, soluble monomer), or with 1.5 × 106 native (NM) or cholesterol-depleted (CDM) hRBCs for 30 min at 37°C. The samples were solubilized with SDS sample buffer, the monomer and oligomeric complexes were resolved on a 1.5% SDS/AGE gel and immunoblotted, and the toxin was detected with specific antibody. Shown are the autoradiograms of the monomer and oligomer bands recognized by the antibody. Immunoblot analysis of untreated erythrocytes did not reveal any cross-reactive proteins (data not shown).
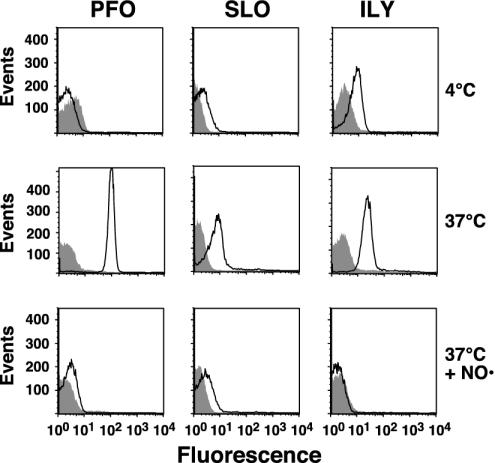
Membrane insertion into native and cholesterol-depleted hRBCs. PFO, SLO, or ILY (9 pmol each), each labeled with NBD in a membrane-facing residue in TMH1, were incubated with 1.5 × 106 hRBCs at 4°C for 30 min and analyzed by flow cytometry. At 4°C, binding and oligomerization proceed, but insertion of the TMHs is significantly inhibited (24). Each sample was then shifted to 37°C for an additional 10 min to allow insertion of the TMHs and then was analyzed again by flow cytometry for changes in NBD emission. To confirm that the increase in NBD intensity seen with native hRBCs was due to TMH membrane insertion, the same sample was equilibrated with the collisional quencher doxyl-stearic acid (37°C + NO·), which places a nitroxide in the bilayer of the erythrocytes. The sample was then analyzed for a third time by flow cytometry. We counted 104 cells per sample in all experiments. Changes in the NBD intensity are shown for the toxins incubated with native (outlined peak) and cholesterol-depleted (shaded peak) hRBCs under each condition.
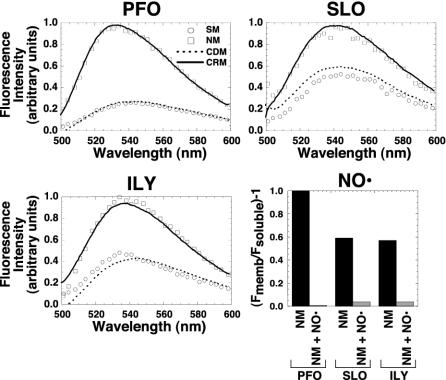
Membrane insertion into native hRBC, cholesterol-depleted, or cholesterol-replete membranes. The fluorescence intensity of the same NBD-labeled toxin used for the experiments in Fig. 4 was determined when they were incubated alone (SM, soluble monomer) or with erythrocyte ghost native membranes (NM), cholesterol-depleted membranes (CDM), or cholesterol-replete membranes (CRP). The samples were incubated for 30 min at 37°C, and the emission intensities of the NBD for each sample were measured between 500 and 600 nm. To confirm that the NBD-labeled residues of SLO and ILY were membrane facing, the same experiment was repeated for each labeled toxin in the absence and presence of the membrane collisional quencher doxyl-stearic acid (graph labeled NO·). Only the net emission intensity of the NBD at 540 nm [(Fmemb/Fsoluble) – 1] is shown for the membrane-inserted toxin on native membranes (NM) vs. membranes containing the doxyl-stearic acid collisional quencher (NM + NO·).
Similar articles
-
A Key Motif in the Cholesterol-Dependent Cytolysins Reveals a Large Family of Related Proteins.mBio. 2020 Sep 29;11(5):e02351-20. doi: 10.1128/mBio.02351-20. mBio. 2020. PMID: 32994330 Free PMC article.
-
The cholesterol-dependent cytolysin family of gram-positive bacterial toxins.Subcell Biochem. 2010;51:551-77. doi: 10.1007/978-90-481-8622-8_20. Subcell Biochem. 2010. PMID: 20213558 Review.
-
Perfringolysin O structure and mechanism of pore formation as a paradigm for cholesterol-dependent cytolysins.Subcell Biochem. 2014;80:63-81. doi: 10.1007/978-94-017-8881-6_5. Subcell Biochem. 2014. PMID: 24798008 Free PMC article. Review.
-
What planar lipid membranes tell us about the pore-forming activity of cholesterol-dependent cytolysins.Biophys Chem. 2013 Dec 1;182:64-70. doi: 10.1016/j.bpc.2013.06.015. Epub 2013 Jul 2. Biophys Chem. 2013. PMID: 23876488 Review.
-
The cholesterol-dependent cytolysins.Curr Top Microbiol Immunol. 2001;257:15-33. doi: 10.1007/978-3-642-56508-3_2. Curr Top Microbiol Immunol. 2001. PMID: 11417120 Review.
Cited by
-
Membrane Binding, Cellular Cholesterol Content and Resealing Capacity Contribute to Epithelial Cell Damage Induced by Suilysin of Streptococcus suis.Pathogens. 2019 Dec 30;9(1):33. doi: 10.3390/pathogens9010033. Pathogens. 2019. PMID: 31905867 Free PMC article.
-
Equinatoxin II permeabilizing activity depends on the presence of sphingomyelin and lipid phase coexistence.Biophys J. 2008 Jul;95(2):691-8. doi: 10.1529/biophysj.108.129981. Epub 2008 Apr 4. Biophys J. 2008. PMID: 18390598 Free PMC article.
-
Perfringolysin O association with ordered lipid domains: implications for transmembrane protein raft affinity.Biophys J. 2010 Nov 17;99(10):3255-63. doi: 10.1016/j.bpj.2010.09.028. Biophys J. 2010. PMID: 21081073 Free PMC article.
-
Intermedilysin cytolytic activity depends on heparan sulfates and membrane composition.PLoS Genet. 2021 Feb 12;17(2):e1009387. doi: 10.1371/journal.pgen.1009387. eCollection 2021 Feb. PLoS Genet. 2021. PMID: 33577603 Free PMC article.
-
The Cholesterol-dependent Cytolysin Membrane-binding Interface Discriminates Lipid Environments of Cholesterol to Support β-Barrel Pore Insertion.J Biol Chem. 2015 Jul 17;290(29):17733-17744. doi: 10.1074/jbc.M115.656769. Epub 2015 Jun 1. J Biol Chem. 2015. PMID: 26032415 Free PMC article.
References
-
- Heuck, A. P., Tweten, R. K. & Johnson, A. E. (2001) Biochemistry 40, 9065–9073. - PubMed
-
- Alouf, J. E. (1999) in Bacterial Toxins: A Comprehensive Sourcebook, eds. Alouf, J. & Freer, J. (Academic, London), pp. 443–456.
-
- Alouf, J. E. (2000) Int. J. Med. Microbiol. 290, 351–356. - PubMed
-
- Jacobs, T., Cima-Cabal, M. D., Darji, A., Méndez, F. J., Vázquez, F., Jacobs, A. A. C., Shimada, Y., Ohno-Iwashita, Y., Weiss, S. & de los Toyos, J. R. (1999) FEBS Lett. 459, 463–466. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
