

Interactions between membrane conductances underlying thalamocortical slow-wave oscillations
- PMID: 14506309
- PMCID: PMC2927823
- DOI: 10.1152/physrev.00012.2003
Interactions between membrane conductances underlying thalamocortical slow-wave oscillations
Abstract
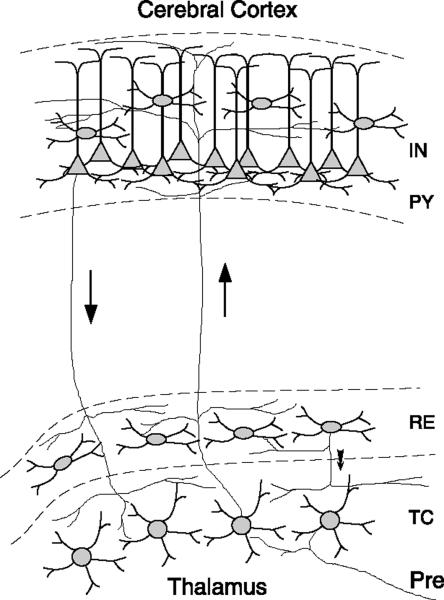
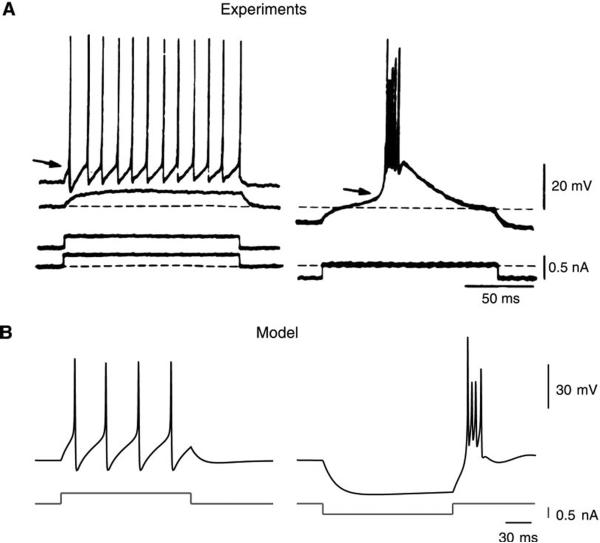
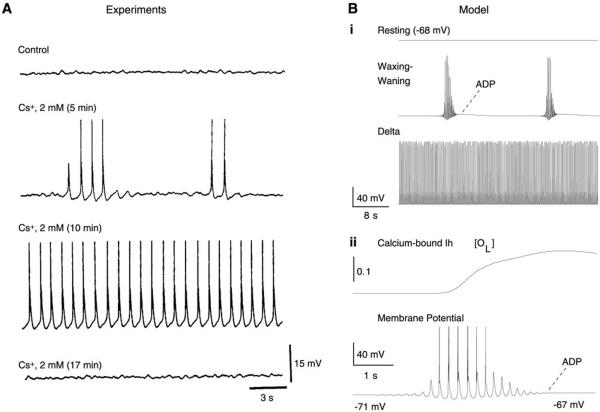
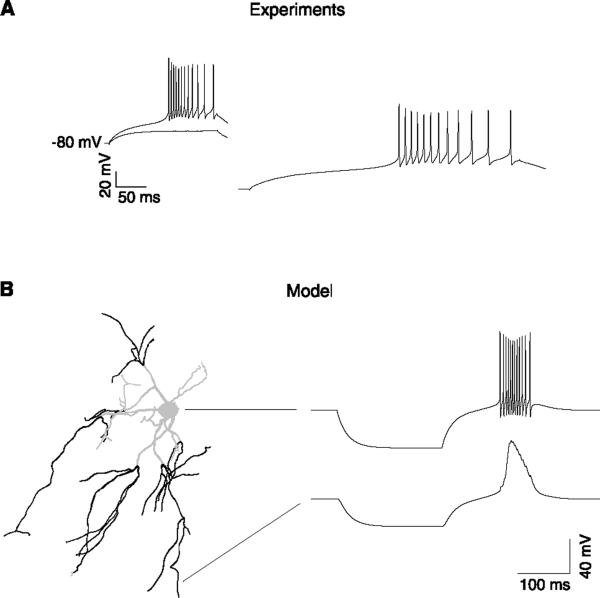
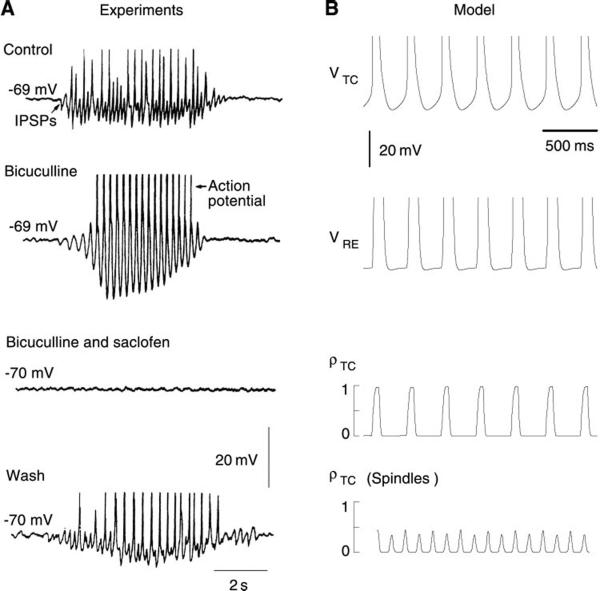
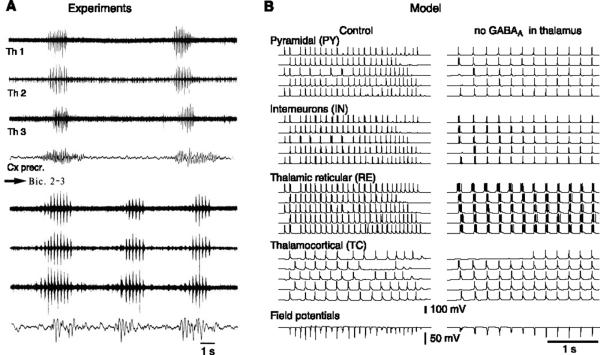
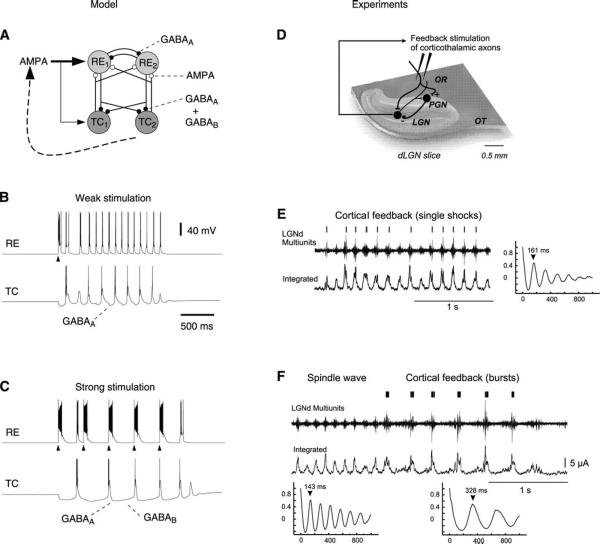
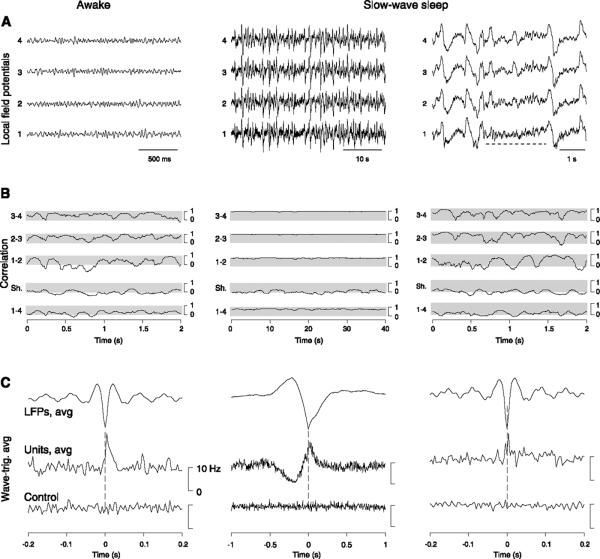
Neurons of the central nervous system display a broad spectrum of intrinsic electrophysiological properties that are absent in the traditional "integrate-and-fire" model. A network of neurons with these properties interacting through synaptic receptors with many time scales can produce complex patterns of activity that cannot be intuitively predicted. Computational methods, tightly linked to experimental data, provide insights into the dynamics of neural networks. We review this approach for the case of bursting neurons of the thalamus, with a focus on thalamic and thalamocortical slow-wave oscillations. At the single-cell level, intrinsic bursting or oscillations can be explained by interactions between calcium- and voltage-dependent channels. At the network level, the genesis of oscillations, their initiation, propagation, termination, and large-scale synchrony can be explained by interactions between neurons with a variety of intrinsic cellular properties through different types of synaptic receptors. These interactions can be altered by neuromodulators, which can dramatically shift the large-scale behavior of the network, and can also be disrupted in many ways, resulting in pathological patterns of activity, such as seizures. We suggest a coherent framework that accounts for a large body of experimental data at the ion-channel, single-cell, and network levels. This framework suggests physiological roles for the highly synchronized oscillations of slow-wave sleep.
Figures
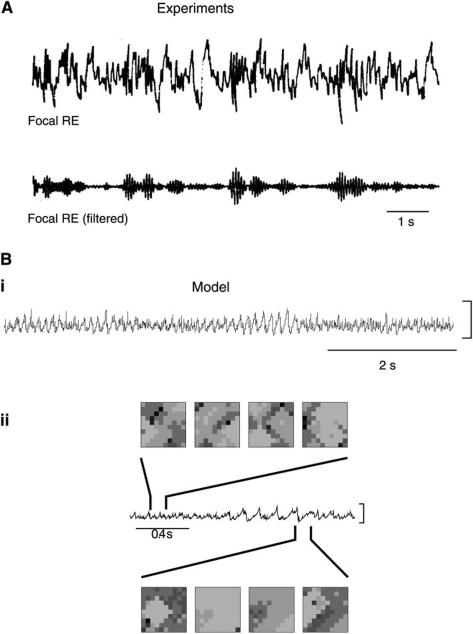

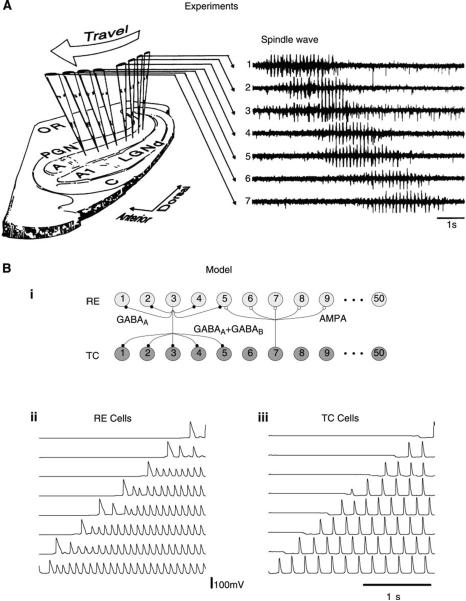
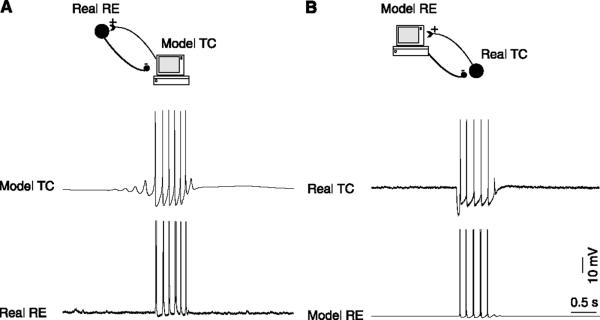
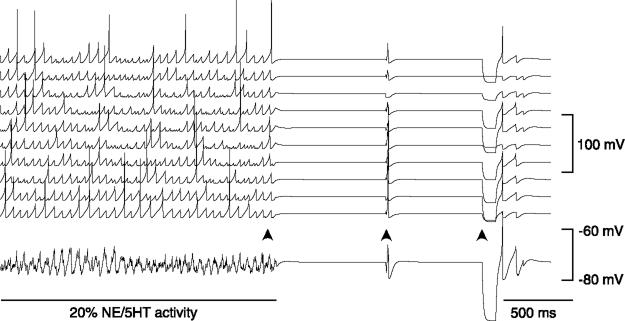
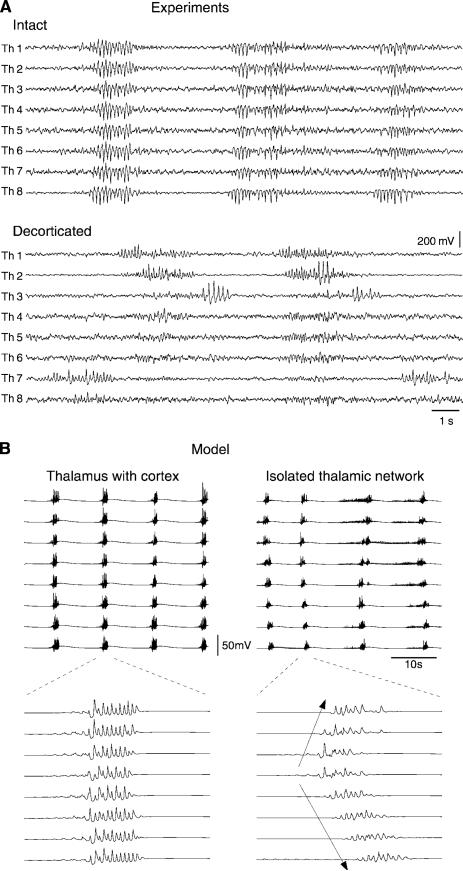
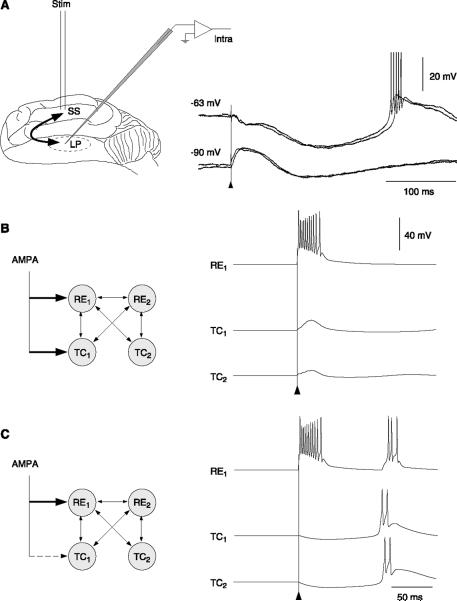
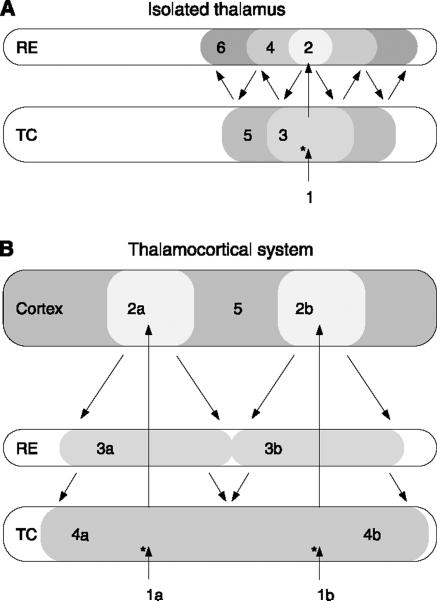
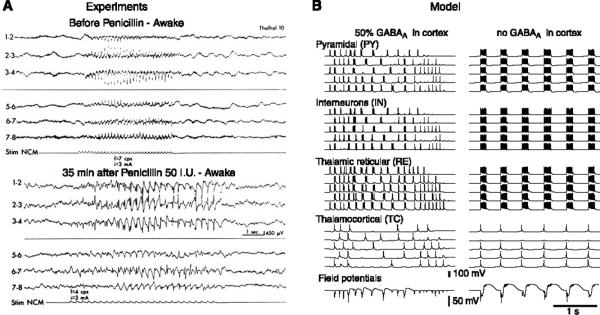
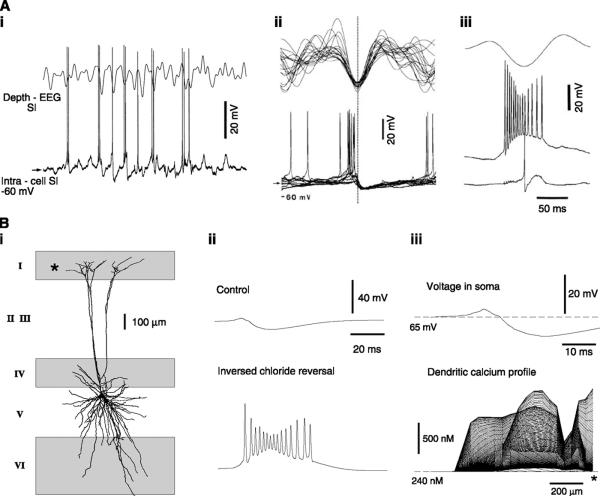
References
-
- Abel T, Nguyen PV, Barad M, Deuel TAS, Kandel ER, Bourtchouladze R. Genetic demonstration of a role for PKA in the late phase of LTP and in hippocampus-based long-term memory. Cell. 1997;88:615–626. - PubMed
-
- Adams DJ, Smith SJ, Thompson SH. Ionic currents in molluscan soma. Annu Rev Neurosci. 1980;3:141–167. - PubMed
-
- Ahlsen G, Grant K, Lindström S. Monosynaptic excitation of principal cells in the lateral geniculate nucleus by corticofugal fibers. Brain Res. 1982;234:454–458. - PubMed
-
- Albowitz B, Kuhnt U. Spread of epileptiform potentials in the neocortical slice: recordings with voltage-sensitive dyes. Brain Res. 1993;631:329–333. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources